- Biology Article
- Pedigree Analysis Family Genetics

Pedigree Analysis - Genetic History of Family
Table of contents, introduction, what is pedigree analysis, pedigree analysis – diagram.
Gregor Mendel worked on the principles of inheritance in genetics a long time ago. His work helped form the fundamentals of genetics and inheritance patterns in living organisms. Later, scientists developed another approach to help us understand the inheritance of genes in living organisms. This approach is known as the pedigree analysis. Let’s now learn about pedigree analysis and how it helps predict genetic diseases.
Pedigree analysis was developed to understand the inheritance of genes from parents to offspring. It was developed as a chart that can represent a family tree along with the family members and their genetic traits, respectively.
Gregor Mendel’s experiments showed that the “factors” that we now know as “genes”, are the factors which are responsible for the inheritance of traits from parents to offspring.
Therefore, genes are the hereditary unit of organisms which are responsible for carrying the information from the parents to their offspring. These genes are responsible for the characteristics of a living organism and can also be the reason for some disorders present in them.
These conclusions are based on the evidence provided by the controlled cross experiments in pea plants and other organisms. Since we can’t perform similar experiments on human populations due to ethical reasons, we formed pedigree analysis to study the pattern of the inheritance even when we have limited data.
Mendelian disorders are genetic diseases which follow a Mendelian inheritance pattern, and their inheritance is controlled by Mendelian genetics. Therefore, by studying the family history of an individual through pedigree analysis, we can predict whether an individual is at risk of a genetic disorder.
A family tree can be represented by a pedigree chart with all the members of a family. They may be having a genetic disorder or maybe a carrier of the disease. In the pedigree analysis, standard symbols are used to distinguish between different families.
Here is a diagrammatic representation of a pedigree chart.

- Here, the rows represent the generations of a family, squares represent males, and circles represent females. Individuals on the same row belong to the same generation.
- A marriage or mating is represented as a horizontal bar between the square and the circle. Likewise, a double bar between the symbols indicates a consanguineous mating or marriage.
- The offsprings are indicated by a suspending vertical line drawn perpendicular to the horizontal bar or marriage bar.
- The homozygous individuals are represented as fully shaded symbols. Whereas heterozygous individuals are just carriers and are represented by half-shaded symbols.
- A normal or natural genotype is denoted as wild type. Unshaded squares and circles indicate the normal or “unaffected” individuals.
- Usually, the carrier of a sex-linked recessive gene is represented by a black spot in the middle of the symbol.
- Sometimes, a particular affected individual who is being interviewed or who bought the trait to the geneticists is indicated by a pointing arrow. Such members are termed, propositus.
For more details on the Pedigree analysis or any other related topic, login to BYJU’S-The Learning App.
Frequently Asked Questions
What do you mean by a pedigree chart.
A pedigree chart is a diagrammatic flow chart that is prepared to exhibit a person’s hereditary information in an easily readable form. The chart usually displays the members of the family who are affected by a particular genetic trait.
What are the main goals of a pedigree analysis?
Pedigree analysis is done to determine if the mode of inheritance is recessive, dominant, partial dominant, autosomal, mitochondrial or sex-linked. Moreover, it also determines the probability of an individual or offspring being affected in the given cross.

Put your understanding of this concept to test by answering a few MCQs. Click ‘Start Quiz’ to begin!
Select the correct answer and click on the “Finish” button Check your score and answers at the end of the quiz
Visit BYJU’S for all Biology related queries and study materials
Your result is as below
Request OTP on Voice Call
| BIOLOGY Related Links | |
Leave a Comment Cancel reply
Your Mobile number and Email id will not be published. Required fields are marked *
Post My Comment
Register with BYJU'S & Download Free PDFs
Register with byju's & watch live videos.
Pedigree Analysis Chart
A pedigree analysis chart provides an important tool to study the inheritance of genes. It also gives precise information about the family members through diagrammatic representation. Sometimes, it is also known as the “ family or genealogic tree “.
A pedigree chart uses internationally accepted symbols, lines, numbers and letters To illustrate different individuals, their interrelationship, the number of individuals, and the number of generations.
It gives an idea to understand the mode of inheritance of human traits and diseases. This post mainly focuses on the definition, symbols, types, examples and importance of the pedigree chart. You will also get to know the points to read the pedigree chart.
Content: Pedigree Analysis Chart
How to read a pedigree chart, definition of pedigree analysis.
It refers to the genetic representation that depicts the family tree or biological relationship between individuals of the new generation and their ancestors. A pedigree chart also entails information about genetic disorders in the family history . It helps in studying different patterns of inheritance and genetic traits of different individuals.
In other words, we can define that pedigree analysis as an analytical method , displaying data on the heredity of traits and disorders. It uses specific symbols to represent the type of individuals and their relationship with each other.
Pedigree Chart Symbols

An arrow pointing towards the symbol indicates proband individuals or they usually have the disorder of interest. A slash through the symbol represents deceased individuals.
A fully shaded symbol indicates affected individuals . A central dot within a symbol depicts a carrier . Two arrows merging on the top of symbols determine fraternal twins .
A line between the two symbols and two arrows merging at the top represents identical twins . A letter ‘A’ within the symbol and double brace indicates an adopted child .
Besides these, other symbols are also illustrated in the above diagram. You can memorize all the symbols given in the diagram above to analyze the heredity of traits and disorders in the family history.
Types of Pedigree Analysis

- Males and females get affected in the same ratio.
- The genetic traits express themselves in each generation or never skips a generation.
- Two affected parents can produce an unaffected child.
- Unaffected parents produce unaffected offspring.
- Heterozygotes are also affected.
Autosomal recessive pedigree chart : To determine autosomal recessive inheritance, look for the following features:
- Males and females are affected in the same ratio.
- The genetic trait may skip a generation.
- Two affected parents will only produce affected children.
- Unaffected parents may produce affected offspring since they are carriers or heterozygotes.
- Heterozygotes have a normal phenotype.
X-linked dominant pedigree chart : To determine x-linked dominant inheritance, look for the following features:
- Both males and females are likely to be affected.
- The genetic traits may skip a generation.
- The affected father can transfer the disease to all female individuals.
- Male to male disease transmission does not occur.
X-linked recessive pedigree chart : To determine x-linked recessive inheritance, you must keep the given features in your mind:
- Males are relatively more affected than females.
- The genetic trait never skips a generation.
- Only the affected mother can transfer the disease to the male individuals.
- Disease transmission from a father to a son will not occur.
Y-linked recessive pedigree chart : To determine y-linked recessive inheritance, you must keep the following features in your mind:
- Only males are affected.
- Y-chromosome carries the genetic trait.
- Disease transmission occurs from a father to a son.
To read a pedigree chart, you must follow the given protocol:

Determine the mode of inheritance : Look into the chart to identify the inheritance pattern, whether it is autosomal or sex-linked. Males and females are equally affected in autosomal inheritance. Oppositely, males are more likely to be affected in sex or x-linked inheritance.
Determination of traits : Then, see the chart to identify the traits (whether dominant or recessive). Dominant traits will never skip a generation, and one of the parents must carry the trait. Recessive traits may skip a generation, and a parent doesn’t need to carry the trait since they can be heterozygous.
Pedigree Analysis Examples
Let us take some examples to solve the pedigree problems or to identify the type of pedigree.
Autosomal Dominant Pedigree

Starting from the left, you can see unaffected parents have unaffected offspring in the F-3 generation, which is also a dominant trait. In the other two relationships, you can see that the ratio of affected males and females is similar, which indicates an autosomal inheritance.
Autosomal Recessive Pedigree

Now, look into the second family in the F-1 generation. All progenies of the F-2 generation acquire the disease due to the affected mother and father, a feature of recessive trait where two affected parents will always produce affected children.
Then, look into the interrelationship between the individuals of the F-2 generation. Individual number 3 and 4 indicates a couple, and they produced infected male and female child in the F-3 generation. Here, in this pedigree, you can see that the ratio of affected males and females is almost the same, which indicates an autosomal inheritance.
X-Linked Dominant Pedigree

We have already discussed that X-linked dominant traits transfer from an affected father to all daughters, and male to male transmission does not occur. Then, look into the interrelationship between the individuals of the F-2 generation.
Individual number 2 and 3 indicates a couple. Again, recall a dominant trait feature where the affected individuals must have one affected parent. So, this scenario confirms that the pedigree shows the dominant trait, not recessive.
Here, you can see a crisscross pattern between individuals in each generation, indicating a sex-linked inheritance mode in this pedigree chart.
X-Linked Recessive Pedigree

Then, look into the interrelationship between the individuals of the F-2 generation. Individual number 1 and 2 indicates a couple. Both parents are unaffected, but the disease has occurred in the F-3 generation, which shows a recessive pattern.
Then, the affected couple (numbers 5 and 6) transfer the disease to all individuals of the F-3 generation, which is again a feature of the recessive trait. Here, you can see more infected males, indicating a mode of sex-linked recessive inheritance.
Y-Linked Pedigree

The second family in the F-1 generation has an unaffected father, so the disease will not transfer to the male individual of the next generation. Then, look into the interrelationship between individuals of the F-2 generation.
Individual number 3 and 4 indicates a couple, where an affected father transferred the disease to his sons in the F-3 generation. You can only see the affected males in this pedigree, which indicates a sex-linked inheritance.
Pedigree analysis reveals the family history by studying the inheritance of traits and diseases between family members across generations. It also helps in genetic counselling . Besides, it determines the dominant or recessive nature of the trait .
The pedigree chart also entails the type of chromosome (autosomal or sex). We can also identify genotypes as well as phenotypes of family members. Determining the probabilities of phenotypes in future generations becomes easy through pedigree analysis.
Related Topics:
- Muscular Tissue
- Qualitative Analysis of Amino Acids
- Cytoplasmic Inheritance
- Gluconeogenesis
- Human Skin Layers
Leave a Comment Cancel Reply
Your email address will not be published. Required fields are marked *
Start typing and press enter to search

Want to create or adapt books like this? Learn more about how Pressbooks supports open publishing practices.
20.3 Pedigree Analysis
What is a pedigree.
A pedigree chart is a diagram that models the inheritance of phenotypes from one generation to the next. Pedigree charts are used by scientists, genetic counselors, and animal breeders. Pedigrees show inheritance across one or more generations. Individuals connected by a horizontal line have mated and had children. Vertical lines connect parents to their children. Siblings are generally shown from left to right according to birth order.

In a pedigree chart, females are symbolized by a circle and males are symbolized by a square. Sometimes the gender of an individual is not identified in the pedigree; these individuals are symbolized by a diamond. A gender may not be specified in the pedigree for one of several reasons: the person’s gender may be unknown, the person may not identify as male or female, the person may be intersex, or the person’s gender may be withheld for privacy reasons. Sometimes diamonds are used for all individuals in a pedigree, when it is not clinically relevant to specify their gender.

Shading represents that the individual is affected by the disease or condition. Pedigree charts do not always include shaded shapes, because sometimes a pedigree is constructed for a reason other than following a disease or condition. Sometimes heterozygotes are indicated by shading half of the shape, but often they are not indicated and are simply shown as unaffected.
Pedigrees can be used to study the genetics of inherited diseases. They can be analyzed to determine whether a genetic condition is autosomal or sex-linked , and whether it is dominant or recessive . We can also ask questions about the genotype of individuals on a pedigree given their phenotypes.
Pedigree Analysis
Pedigree charts display different patterns depending on which inheritance mechanism is responsible for the trait. Scientists can analyze pedigree charts to study how inherited disorders are transmitted. The most common modes of inheritance are autosomal recessive, autosomal dominant, X-linked recessive, and X-linked dominant. There are also a handful of Y-linked and mitochondrial disorders.
Autosomal Recessive Inheritance
Some genetic conditions are autosomal recessive, meaning that the gene involved is found on an autosome, and affected individuals have two copies of the allele that causes the condition. If an affected individual in a pedigree has two unaffected parents, the condition is most likely recessive. Additionally, if daughters in the pedigree have two unaffected parents, the condition is most likely autosomal recessive (unlike X-linked recessive conditions, in which an affected daughter will have an affected father). With autosomal recessive inheritance, males and females are equally likely to be affected.
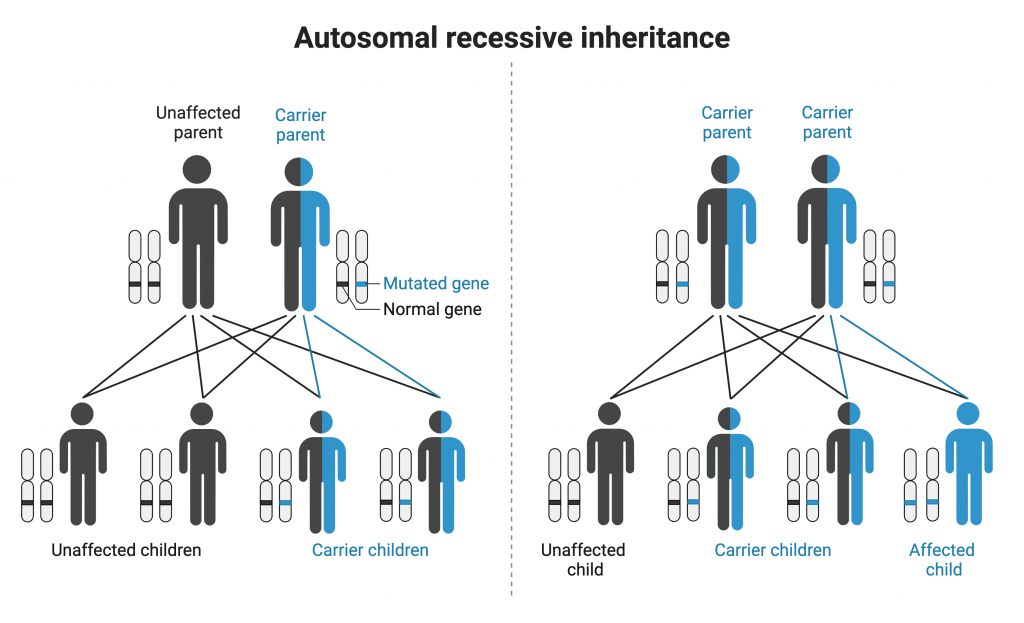
Autosomal recessive conditions in humans include cystic fibrosis, sickle cell disease, Tay-Sachs disease, and phenylketonuria.

It is helpful to be able to assign genotypes to all individuals on a pedigree chart in order to predict if it is possible that an individual might pass along a mutant allele to their offspring. When the inheritance mechanism is autosomal recessive, it is helpful to start with affected individuals. All affected individuals are expected to be homozygous recessive in genotype (for example, genotype aa). Looking at individual 7 on the pedigree chart above, we would then recognize that both parents (individuals 1 and 2) must carry a copy of the mutant allele. Both parents are expected to be heterozygous in genotype because neither one is affected by the trait. One can then determine the genotypes of the siblings of individual 7. Here, we do not know whether individuals 5 and 6 are heterozygous or homozygous for the normal allele because no offspring are shown. However, we can conclude that individual 4 is heterozygous for the relevant gene because her daughter (individual 10) expresses the trait. Furthermore, one can conclude that both individual 3 and individual 11 must also be heterozygous for the relevant gene.
Autosomal Dominant Inheritance
Genetic conditions can display autosomal dominance. In this mode of transmission, a single mutant allele is sufficient to cause the condition because the mutant allele is dominant over the normal allele. Affected children generally have at least one affected parent (although not always, because some conditions display incomplete penetrance, meaning that not every individual with the allele will display the phenotype). Males and females are equally affected.

Autosomal dominant conditions in humans include Marfan syndrome, Huntington’s disease, and achondroplasia.

Let’s think about assigning genotypes to individuals on a pedigree chart representing an autosomal dominant trait. Each affected individual must have one or two copies of the mutant allele; each unaffected individual is expected to be homozygous for the normal allele (genotype aa). One of the 4 children of parents 1 and 2 is not affected by the trait and hence must have two copies of the normal allele which acts in a recessive fashion (genotype aa). This would suggest that individual 1 is heterozygous in genotype (Aa). Furthermore, individuals 6, 8, and 9 are also expected to be heterozygous because their father is homozygous recessive in genotype.
X-linked Recessive Inheritance
Some conditions are sex-linked, meaning the gene that causes the condition is on one of the sex chromosomes. In mammals, this is usually the X chromosome, because the X chromosome is much larger and has many more genes. X-linked recessive conditions are much more common in males, because they have only one X chromosome. Therefore, if they inherit an X chromosome with the mutated allele that causes the condition, they will display the condition. Females must inherit two copies to display an X-linked recessive condition. Males cannot be carriers; they will either be affected or unaffected, depending on which allele they inherit on the X chromosome.

In pedigrees that display X-linked recessive inheritance, sons of a carrier mother and an unaffected father have a 50% chance of being affected, while daughters will not be affected, but have a 50% chance of being carriers (heterozygotes). Sons of an affected mother will all be affected. There is no father-to-son transmission of X-linked traits.
Examples of X-linked recessive conditions include Duchenne muscular dystrophy, the most common form of color blindness, and hemophilia A and B.
To assign genotypes to individuals on a pedigree chart representing an X-linked recessive trait, it is important to recognize two things: 1. Males only have one copy of the X chromosome and females have two copies, and 2) the presence of one normal allele will result in an unaffected individual. Thus all unaffected males will have a copy of the normal allele (genotype X A Y, and all affected males will have a copy of the mutant allele (genotype X a Y). All three of the affected males are expected to have the X a Y genotype, having received the X chromosome from their mother. Thus individuals 1 and 7 are both heterozygous females with genotype X A X a .
X-linked Dominant Inheritance
Conditions that display X-linked dominant inheritance are less common than X-linked recessive conditions. For this mode of inheritance, affected fathers will pass on the allele and condition to all of their daughters. Sons of an affected, heterozygous mother have a 50% chance of being affected.
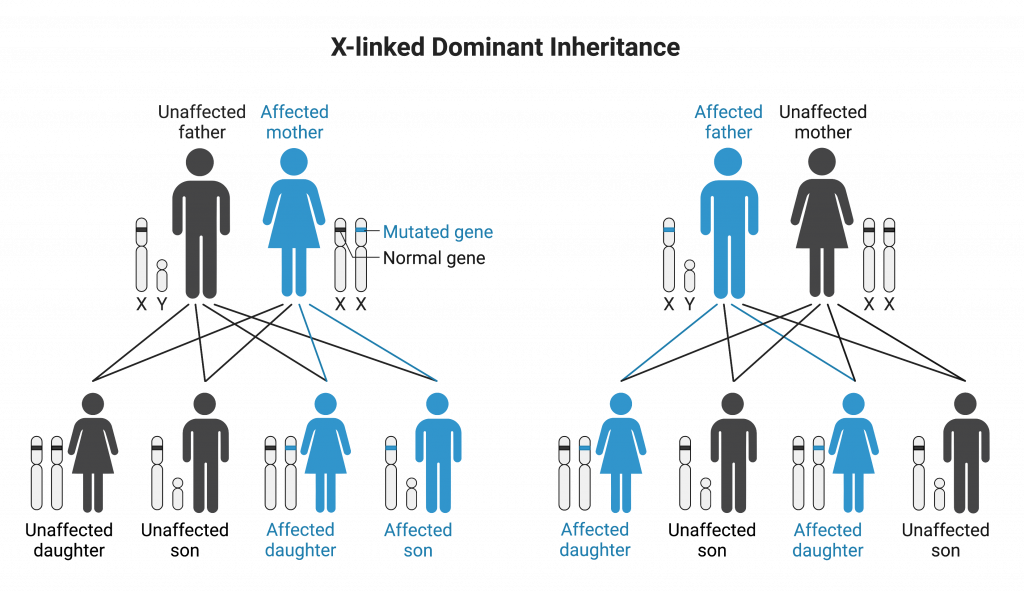
Examples of X-linked dominant inheritance include Rett syndrome and most cases of Alport syndrome.

In order for a daughter to be homozygous for the mutant allele, both of her parents must be affected by the mutant trait. Thus all affected females on this chart are heterozygous in genotype.
trait or condition that is transmitted on one of the autosomes (chromosomes that are not sex chromosomes)
trait or condition transmitted on one of the sex chromosomes (in mammals, usually the X chromosome)
trait which confers the same physical appearance whether an individual has two copies of the trait or one copy of the dominant trait and one copy of the recessive trait
trait that appears “latent” or non-expressed when the individual also carries a dominant trait for that same characteristic; when present as two identical copies, the recessive trait is expressed
College Biology I Copyright © by Melissa Hardy is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License , except where otherwise noted.

- school Campus Bookshelves
- menu_book Bookshelves
- perm_media Learning Objects
- login Login
- how_to_reg Request Instructor Account
- hub Instructor Commons
Margin Size
- Download Page (PDF)
- Download Full Book (PDF)
- Periodic Table
- Physics Constants
- Scientific Calculator
- Reference & Cite
- Tools expand_more
- Readability
selected template will load here
This action is not available.
5.2: Pedigree Analysis
- Last updated
- Save as PDF
- Page ID 4036

- Todd Nickle and Isabelle Barrette-Ng
- Mount Royal University & University of Calgary
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
Pedigree charts are diagrams that show the phenotypes and/or genotypes for a particular organism and its ancestors. While commonly used in human families to track genetic diseases, they can be used for any species and any inherited trait. Geneticists use a standardized set of symbols to represent an individual’s sex, family relationships and phenotype. These diagrams are used to determine the mode of inheritance of a particular disease or trait, and to predict the probability of its appearance among offspring. Pedigree analysis is therefore an important tool in both basic research and genetic counseling .
Each pedigree chart represents all of the available information about the inheritance of a single trait (most often a disease) within a family. The pedigree chart is therefore drawn using factual information, but there is always some possibility of errors in this information, especially when relying on family members’ recollections or even clinical diagnoses. In real pedigrees, further complications can arise due to incomplete penetrance (including age of onset) and variable expressivity of disease alleles, but for the examples presented in this book, we will presume complete accuracy of the pedigrees. A pedigree may be drawn when trying to determine the nature of a newly discovered disease, or when an individual with a family history of a disease wants to know the probability of passing the disease on to their children. In either case, a tree is drawn, as shown in Figure \(\PageIndex{2}\), with circles to represent females, and squares to represent males. Matings are drawn as a line joining a male and female, while a consanguineous mating (closely related is two lines.

The affected individual that brings the family to the attention of a geneticist is called the proband (or propositus). If an individual is known to have symptoms of the disease ( affected ), the symbol is filled in. Sometimes a half-filled in symbol is used to indicate a known carrier of a disease; this is someone who does not have any symptoms of the disease, but who passed the disease on to subsequent generations because they are a heterozygote. Note that when a pedigree is constructed, it is often unknown whether a particular individual is a carrier or not, so not all carriers are always explicitly indicated in a pedigree. For simplicity, in this chapter we will assume that the pedigrees presented are accurate, and represent fully penetrant traits.
If you're seeing this message, it means we're having trouble loading external resources on our website.
If you're behind a web filter, please make sure that the domains *.kastatic.org and *.kasandbox.org are unblocked.
To log in and use all the features of Khan Academy, please enable JavaScript in your browser.
AP®︎/College Biology
Course: ap®︎/college biology > unit 5.
- Variations on Mendel's laws (overview)
- Thomas Hunt Morgan and fruit flies
- The chromosomal basis of inheritance
- X-linked inheritance
- Genetic linkage & mapping
- Pedigree for determining probability of exhibiting sex linked recessive trait
Pedigrees review
- Extranuclear inheritance 1
- Inheritance of mitochondrial and chloroplast DNA
- Non-Mendelian genetics
| Term | Meaning |
|---|---|
| Pedigree | Chart that shows the presence or absence of a trait within a family across generations |
| Genotype | The genetic makeup of an organism (ex: TT) |
| Phenotype | The physical characteristics of an organism (ex: tall) |
| Dominant allele | Allele that is phenotypically expressed over another allele |
| Recessive allele | Allele that is only expressed in absence of a dominant allele |
| Autosomal trait | Trait that is located on an autosome (non-sex chromosome) |
| Sex-linked trait | Trait that is located on one of the two sex chromosomes |
| Homozygous | Having two identical alleles for a particular gene |
| Heterozygous | Having two different alleles for a particular gene |
Reading a pedigree
- Determine whether the trait is dominant or recessive. If the trait is dominant, one of the parents must have the trait. Dominant traits will not skip a generation. If the trait is recessive, neither parent is required to have the trait since they can be heterozygous.
- Determine if the chart shows an autosomal or sex-linked (usually X-linked) trait. For example, in X-linked recessive traits, males are much more commonly affected than females. In autosomal traits, both males and females are equally likely to be affected (usually in equal proportions).
Example: Autosomal dominant trait
Example: x-linked recessive trait, common mistakes and misconceptions.
- The presence of many affected individuals in a family does not always mean that the trait is dominant. The terms dominant and recessive refer to the way that a trait is expressed, not by how often it shows up in a family. In fact, although it is uncommon, a trait may be recessive but still show up in all generations of a pedigree.
- You may not always be able to determine the genotype of an individual based on a pedigree. Sometimes an individual can either be homozygous dominant or heterozygous for a trait. Often, we can use the relationships between an individual and their parents, siblings, and offspring to determine genotypes. However, not all carriers are always explicitly indicated in a pedigree, and it may not be possible to determine based on the information provided.
Want to join the conversation?
- Upvote Button navigates to signup page
- Downvote Button navigates to signup page
- Flag Button navigates to signup page


Want to create or adapt books like this? Learn more about how Pressbooks supports open publishing practices.
11 Chapter 11: Pedigrees
Lisa Limeri; Shifath Bin Syed; Joshua Reid; and rocksher
Learning Objectives
By the end of this section, you will be able to do the following:
- Using a pedigree for an autosomal or sex-linked trait, determine the genotype and phenotype for each person in the pedigree.
- Given information on parental and offspring phenotypes, determine whether the alleles involved are 1) dominant, recessive, or codominant, 2) autosomal or X-linked, and 3) linked or unlinked.
Introduction
Many human diseases are genetically inherited. A healthy person in a family in which some members suffer from a recessive genetic disorder may want to know if he or she has the disease-causing gene and what risk exists of passing the disorder on to his or her offspring. Of course, doing a test cross in humans is unethical and impractical. Instead, geneticists use pedigree analysis to study the inheritance pattern of human genetic diseases.
Pedigree Analysis
Pedigree charts show the phenotypes and/or genotypes of a particular organism and its ancestors. While commonly used in human families to track genetic diseases, they can be used for any species and inherited trait. Geneticists use standardized symbols to represent an individual’s sex, family relationships, and phenotype. These diagrams are used to determine the mode of inheritance of a particular disease or trait and to predict the probability of its appearance among offspring. Therefore, Pedigree analysis is an important tool in basic research and genetic counseling.
Each pedigree chart represents all available information about the inheritance of a single trait (most often a disease) within a family. The pedigree chart is therefore drawn using factual information. Still, there is always some possibility of errors in this information, especially when relying on family members’ recollections or even clinical diagnoses. In real pedigrees, further complications can arise due to incomplete penetrance (including the age of onset) and variable expressivity of disease alleles. Despite these real-world complications, for the examples presented in this book, we will assume complete accuracy of the pedigrees. A pedigree may be drawn when determining the nature of a newly discovered disease or when an individual with a family history of a disease wants to know the probability of passing the disease on to their children. In either case, a tree is drawn, as shown in Figure 11.1, with circles representing females and squares representing males. Matings are drawn as a line joining a male and female, while consanguineous mating (mating between closely related individuals) is two lines.

The affected individual that brings the family to the attention of a geneticist is called the proband. If an individual is known to have symptoms of the disease ( affected ), the symbol is filled in. Sometimes a half-filled-in symbol is used to indicate a known carrier of a disease; this is someone who does not have any symptoms of the disease but who passed the disease on to subsequent generations because they are a heterozygote. Note that when a pedigree is constructed, it is often unknown whether a particular individual is a carrier or not, so not all carriers are always explicitly indicated in a pedigree.

Reading Question #1
In a pedigree chart, what does a half-filled-in symbol represent?
A. An affected individual with symptoms of the disease.
B. An unaffected individual without any disease-causing allele.
C. An individual who is a known carrier of a disease.
D. An individual with an unknown genotype.
Reading Question #2
What is the purpose of pedigree analysis?
A. To determine the mode of inheritance of a genetic disease or trait.
B. To perform test crosses in humans to identify disease-causing genes.
C. To draw diagrams representing the phenotype and genotype of an organism.
D. To accurately track the family history of genetic diseases.
Inferring the Mode of Inheritance
Given a pedigree of an uncharacterized disease or trait, one of the first tasks is to determine which inheritance modes are possible and which mode of inheritance is most likely. This information is essential in calculating the probability that the trait will be inherited in any future offspring. We will mostly consider five major types of inheritance: autosomal dominant, autosomal recessive, X-linked dominant, X-linked recessive, and Y-linked.
Autosomal Dominant
When a dominant allele of a gene causes a disease, every person with that allele will show symptoms of the disease (assuming complete penetrance), and only one disease allele needs to be inherited for an individual to be affected. Thus, every affected individual must have an affected parent. A pedigree with affected individuals in every generation is typical of autosomal dominant diseases. However, beware that other modes of inheritance can also show the disease in every generation, as described below. It is also possible for an affected individual with an autosomal dominant disease to have a family without any affected children if the affected parent is a heterozygote. This is particularly true in small families, where the probability of every child inheriting the normal, rather than disease allele, is not extremely small. Note that autosomal dominant diseases are usually rare in populations; therefore, affected individuals with autosomal dominant diseases tend to be heterozygotes (otherwise, both parents would have had to be affected with the same rare disease). Achondroplastic dwarfism and polydactyly are examples of human conditions that may follow an autosomal dominant mode of inheritance.

X-linked Dominant
In X-linked dominant inheritance, the gene responsible for the disease is located on the X-chromosome, and the allele that causes the disease is dominant to the normal allele in females. Because females have 2 X-chromosomes whereas males have only 1, females tend to be more frequently affected than males in the population. However, not all pedigrees provide sufficient information to distinguish x-linked dominant and autosomal dominant. One definitive indication that a trait is inherited as autosomal dominant and not X-linked dominant, is when an affected father passes the disease to a son. This type of transmission is not possible with X-linked dominant inheritance since males inherit their X chromosome from their mothers.
Autosomal Recessive
Diseases inherited in an autosomal recessive pattern require that both parents of an affected individual carry at least one copy of the recessive disease allele. With autosomal recessive traits, many individuals in a pedigree can be carriers, probably without knowing it. Compared to pedigrees of dominant traits, autosomal recessive pedigrees tend to show fewer affected individuals and are more likely than autosomal dominant or X-linked dominant to “skip a generation.” Thus, the major feature distinguishing autosomal recessive from dominant is that unaffected individuals can have affected offspring.
Many inborn errors of metabolism, such as phenylketonuria (PKU), are inherited as autosomal recessive. Newborns are often tested for a few of the most common metabolic diseases. A pedigree consistent with autosomal recessive inheritance is displayed in Figure 11.4 below.

Everyday Connection: Carrier Screening
Carrier screening for autosomal recessive disorders is a “well-established public health measure that has substantially decreased the burden of a variety of diseases, particularly in individuals belonging to certain genetic risk groups or with a family history of genetic disease. Notable examples of the success of such screening programs when combined with pretest and posttest genetic counseling include the reduction of β-thalassemia cases in Greece and Italy by 80–90% and the diminishing of Tay–Sachs disease by ~90% in Ashkenazi Jews in the US and Israel” (Xiao & Lauschke, 2021).
As carrier screening methods improve, analyses of genetic disease structures in smaller geographic areas will be refined. This is important “for isolated populations, in which founder mutations can cause high population-specific prevalence of genetic conditions and diseases, as shown for Inuit, and Druze communities in Israel, and Dutch and Finnish populations. These founder mutations are often not covered in conventional testing panels and, consequently, members of these isolated populations might not optimally benefit from pan-ethnic risk variant profiling” (Xiao & Lauschke, 2021).
X-linked Recessive
Because males have only one X-chromosome, any male that inherits an X-linked recessive disease allele will be affected by it. Therefore, in X-linked recessive mode of inheritance, males tend to be affected more frequently than females in a population. This is in contrast to autosomal traits, where both sexes tend to be affected equally, and X-linked dominant, where females are affected more frequently. Note, however, in the small sample sizes typical of human families, it is usually not possible to accurately determine whether one sex is affected more frequently than others. On the other hand, one feature of a pedigree that can be used to definitively establish that an inheritance pattern is not X-linked recessive is the presence of an affected daughter from unaffected parents; because she would have had to inherit one X-chromosome from her father, he would also have been affected in X-linked recessive.
Some forms of color blindness are inherited as X-linked recessive traits. A pedigree consistent with X-linked recessive inheritance is displayed in Figure 11.5 below.

Y-linked and Mitochondrial Inheritance
Two additional modes are Y-linked and Mitochondrial inheritance .
Only males are affected in human Y-linked inheritance (and other species with the X/Y sex-determining system). There is only father-to-son transmission. This is the easiest mode of inheritance to identify, but it is one of the rarest because there are so few genes located on the Y-chromosome. An example of Y-linked inheritance is the hairy-ear-rim phenotype. As expected, this trait is passed on from father to all sons and no daughters. Y-chromosome DNA polymorphisms can be used to follow the male lineage in large families or through ancient ancestral lineages. For example, the Y-chromosome of Mongolian ruler Genghis Khan, and his male relatives, account for ~8% of the Y-chromosome lineage of men in Asia (0.5% worldwide).
Mutations in Mitochondrial DNA (mtDNA) are inherited through the maternal line. There are some human diseases associated with mutations in mitochondrial genes. These mutations can affect both males and females, but males cannot pass them on as the mitochondria are inherited via the egg, not the sperm. Mitochondrial DNA polymorphisms are also used to investigate evolutionary lineages, both ancient and recent. Because of the relative similarity of sequence, mtDNA is also used in species identification in ecology studies.
Reading Question #3
Which mode of inheritance is characterized by the presence of affected individuals in every generation?
A. Autosomal dominant
B. X-linked dominant
C. Autosomal recessive
D. X-linked recessive
Reading Question #4
Which mode of inheritance is characterized by the transmission of traits from fathers to all sons but no daughters?
D. Y-linked
Reading Question #5
A pedigree shows a disease that affects both males and females, with unaffected parents having affected offspring. Which mode of inheritance is most likely?
A) Autosomal dominant
B) X-linked dominant
C) Autosomal recessive
D) X-linked recessive
References and Acknowledgements
Xiao, Q. and Lauschke, V.M. (2021). The prevalence, genetic complexity, and population-specific founder effects of human autosomal recessive disorders. npj Genom . Med. 6 (41). doi: 10.1038/s41525-021-00203-x
Adapted from Nickle T. and Barrette-Ng, I. (2022). 5: Pedigrees and Populations. Online Open Genetics. LibreTexts . Retrieved from https://bio.libretexts.org/Bookshelves/Genetics/Online_Open_Genetics_(Nickle_and_Barrette-Ng)/05%3A_Pedigrees_and_Populations
Introductory Biology I Copyright © by Lisa Limeri; Shifath Bin Syed; Joshua Reid; and rocksher is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License , except where otherwise noted.
Share This Book
Browse Course Material
Course info, instructors.
- Prof. Eric Lander
- Prof. Robert Weinberg
- Prof. Tyler Jacks
- Prof. Hazel Sive
- Prof. Graham Walker
- Prof. Sallie Chisholm
- Dr. Michelle Mischke
Departments
As taught in, learning resource types, fundamentals of biology.
« Previous | Next »
Session Overview
| Genetics in humans cannot be studied by performing controlled crosses rather, analysis of inheritance patterns in an existing population must be used. An approach, called pedigree analysis, is used to study the inheritance of genes in humans. This session will outline how to construct a family pedigree, and how to interpret the information in a pedigree using Mendel’s laws of inheritance and an understanding of the chromosome theory of inheritance. . . |
Session Activities
Lecture video.
For this session, watch the video lecture called “Human Genetics” by Prof. Eric Lander recorded in 2004, starting partway into the lecture until the end.
- Human Genetics
Check Yourself
The following pedigree represents the inheritance pattern of a specific genetic trait in humans. Assume the trait is common in the population.  This disorder could be inherited in which of the following ways? as an Autosomal Dominant trait close as an Autosomal Recessive trait close as an X-linked Dominant trait close as an X-linked recessive trait close as both an Autosomal Dominant trait and an X-linked Dominant trait close as both an Autosomal Recessive trait and an X-linked recessive trait check Check
Help Session Video
Watch the short video of Genny Gould explaining how to analyze a human pedigree (Question 2 in the practice problems below).
Practice Problems
- Problems (PDF)
- Solutions (PDF)
Further Study
Suggested topics for further study in an introductory-level biology textbook.
- Constructing a family pedigree
- Pedigree analysis
- Chromosome theory of inheritance
Useful Links
- Pedigree Analysis, Example 1
- Pedigree Analysis, Example 2
- Pedigree Analysis, Example 3

You are leaving MIT OpenCourseWare

Snapsolve any problem by taking a picture. Try it in the Numerade app?
Find Study Materials for
- Explanations
- Business Studies
- Combined Science
- Computer Science
- Engineering
- English literature
- Environmental Science
- Human Geography
- Macroeconomics
- Microeconomics
- Social Studies
- Browse all subjects
- Textbook Solutions
- Read our Magazine
Create Study Materials
- Flashcards Create and find the best flashcards.
- Notes Create notes faster than ever before.
- Study Sets Everything you need for your studies in one place.
- Study Plans Stop procrastinating with our smart planner features.
- Pedigree Analysis
Pedigrees are some of the most fun and exciting tools we have in inheritance studies. Learning how to analyze them requires pattern recognition and deductive reasoning, but these learning processes are not complicated since they are visual. By using common sense and some fundamental principles, we can analyze pedigrees for just about any trait - from black hair color to osteogenesis imperfecta to dimples.

Create learning materials about Pedigree Analysis with our free learning app!
- Instand access to millions of learning materials
- Flashcards, notes, mock-exams and more
- Everything you need to ace your exams
- Bioenergetics
- Biological Molecules
- Biological Organisms
- Biological Processes
- Biological Structures
- Biology Experiments
- Cell Communication
- Cellular Energetics
- Chemistry of Life
- Communicable Diseases
- Control of Gene Expression
- Ecological Levels
- Energy Transfers
- Genetic Information
- Allopatric Speciation
- Biological Species Concept
- Biotic and Abiotic Factors
- Causes Of Extinction
- Chromosomal Disorders
- Chromosomal Theory of Inheritance
- Common Ancestry
- Comparison of Mitosis and Meiosis
- Darwins Finches
- Ecological Levels of Organization
- Evidence of Evolution
- Evolutionary Changes
- Evolutionary Fitness
- Fossil Record
- Genetic Disorders
- Genetic Drift
- Genetic Recombination
- Genomic Imprinting
- Horizontal Gene Transfer
- Law of Independent Assortment
- Linked Genes
- Mass Extinction
- Mendel's Law of Segregation
- Mendelian Genetics
- Miller Urey Experiment
- Molecular Clock
- Morphological Homology
- Non Nuclear Inheritance
- Oparin-Haldane Hypothesis
- Origin of Life Theories
- Origin of Life on Earth
- Phenotypic Plasticity
- Phenotypic Variations
- Phylogenetic Tree
- Population Genetics
- Probabilities in Genetics
- Punnett Squares
- RNA World Hypothesis
- Reproductive Isolation
- Sex-Linked Traits
- Sympatric Speciation
- Types of Mutations
- Units of Inheritance
- X-linked Genes
- Microbiology
- Organ Systems
- Plant Biology
- Reproduction
- Responding to Change
- Substance Exchange
Definition of Pedigree Analysis in Biology
Pedigree analysis is an examination, or demonstration of the inheritance pattern of particular trait(s) in human beings. It can be represented by a pedigree chart , which is a visual representation of a family tree linking family members and their genetic trail.
In the context of genetics, medicine, and biology, t hese traits are typically diseases and disorders. Think of pedigrees as a family tree, but instead of perhaps going into detail about ethnic backgrounds or country of origin, pedigrees describe who has, who doesn't have, and who carries a genetic disorder (or multiple disorders!).
Pedigree analysis is visualized with a chart or diagram that maps out all relevant members of a family and exactly how they are related to one another. Most pedigrees have a basic level of detail - they demonstrate who's married to who, who is deceased, and the number of progeny and their sex.
Some pedigrees are more detailed, perhaps demonstrating the cause of death for those deceased, or adopted vs biological children.
Regardless of their level of detail, pedigrees highlight who is affected by the disorder in question and who is not. Those affected are typically marked in black, while those unaffected (considered the normal phenotype) have no coloring (or white). The typical denotations in the pedigree analysis are seen below (Fig. 1).
Pedigrees easily demonstrate the phenotypes of the individuals being studied. We can then use them to determine the genotypes of existing family members. We can even use them to predict the genotype and phenotype of future offspring, like in a married couple who wants to know the odds of one of their children having a particular disease.
Pedigree Analysis Examples
We know the basic structures of pedigrees, the meaning of their symbols, and that they are used in genetics to study inheritance patterns. But what are the possible inheritance patterns we can see using pedigrees? And which traits have which kind of particular inheritance pattern? We will determine the answers to these questions with examples of each pedigree, of which there are six in Mendelian genetics.
1. Autosomal Recessive Trait - a Pedigree Analysis of Galactosemia
The first and most common inheritance pattern that can be analyzed by pedigree is that of the autosomal recessive trait . What kind of genes have an autosomal recessive pattern? Thankfully, most genetic diseases! Why thankfully? Well, because an autosomal recessive trait must have two alleles in order for it to appear in the phenotype of an individual, thus the chances of suffering from a recessive trait are lower than those of suffering from a dominant trait. This creates three classes of individuals when it comes to an autosomal recessive pattern of inheritance: those who have the disease ( homozygous recessive ), those who are carriers ( heterozygous ), and those who are neither ( homozygous dominant ).
Let's use the disease galactosemia to study this.
Galactosemia is a disorder of galactose accumulation in the blood due to a defect in the enzyme that metabolizes it.
This build-up of galactose can be toxic to certain tissues of the body. Galactose is present in lactose, which is present in milk, so the first symptoms of galactosemia usually appear in the first couple days of life, after the new baby drinks formula or breast milk. These symptoms include vomiting, diarrhea, being really weak, and even cataracts due to excess galactose in the eyes.
Galactosemia: galacto - referring to galactose, a sugar, semia - in the blood.
Galactosemia is an autosomal recessive disorder. Both mom and dad need at least one copy of this disorder for one of their children to have it. Let's look at a pedigree with such a scenario (Fig. 2).
This is a simple pedigree, but we can see that this heterozygous couple (genotypes Gg) had one child with galactosemia, and three children with the normal phenotype. Because this is an autosomal recessive trait, carriers will not have the disease or any symptoms.
What if we were look at a pedigree analysis of galactosemia (or any other autosomal recessive trait), but it was not labelled as such? What tricks would we use to classify the trait being studied in the pedigree as autosomal recessive? Let's look at an unlabeled example to assess this (Fig. 3).
L ooking at the youngest generation (generations are often labelled, with the oldest generation being I , their descendants being II , and the youngest being III ), we can see there is a male child who is affected by galactosemia. We can see that neither his sisters nor his parents have galactosemia. Thus, we can rule out:
- autosomal dominant (at least one parent would have to have the disorder to pass it down),
- Y-linked (the father must pass this down, so he would have the disorder)
- mitochondrial (a mother passes this down to all her children), and
- X-linked dominant (the mother would have to have this disorder to pass it down to her son). However, we cannot rule out X-linked recessive.
Let us look further up this pedigree, at the first generation. We see there is another person affected with this trait in this family, in Generation-I. This affected woman gave birth to two sons, and two daughters. None of those children are affected (draw this pedigree yourself to get some practice!).
Let us consider the scenario in which the trait this woman has is X-linked recessive. She would have to have two copies of this allele to have the disease, because women have two X-chromosomes. So she would only have the disordered allele to give to her children, and while her daughters would get a normal X-chromosome from their father, all her sons would get an affected X-chromosome from her and a normal Y-chromosome from their father, and thus would have to be affected as well. That is not the inheritance pattern we see here, and this rules out the possibility that this trait, in this case galactosemia, is inherited in an X-linked recessive pattern.
Now, if this affected woman in Generation-I has an autosomal recessive trait, then her genotype must include two copies of the affected allele ( gg ) and she would once again be able to distribute only this to her offspring, However, if their father has a homozygous healthy genotype, GG (which is typically assumed), then all their progeny would have the Gg heterozygous genotype. All four of their children in Generation-II would be carriers . This explains how the woman in Generation-II, who married a random man who perhaps was a carrier as well, gave rise to an affected offspring.
2. Autosomal Dominant - How We Can Use Pedigree Analysis to Find Genetic Disorders With This Inheritance Pattern
Generally, autosomal dominant disorders are present in every generation. This is in contrast to autosomal recessive disorders that are said to "skip generations". Autosomal dominant traits are one of the easiest to recognize on pedigrees because every person exhibiting the trait has at least one parent exhibiting the trait. (Fig. 4)
Let's say this is a pedigree of a family with Huntington's, a disease that causes problems with movement, neurological and psychiatric problems, often resulting in premature death. How can we know that this disease is inherited in an autosomal dominant fashion? We see in Generation-I an affected man passes it on to three of his children- two daughters and one son. Each affected person in Generation-II passes the disorder on to at least one of their children, and the Generation-II son who did not inherit the disorder, and did get married, did not pass it on to any of his four children. Thus, that son is homozygous for the normal allele, and the affected individuals are all heterozygous for this trait.
3. X-linked Recessive
X-linked recessive disorde rs are passed from a woman (who is typically a heterozygote carrier) to both her sons and daughters. However, all her sons will have the trait of the disorder, and her daughters (assuming her husband has the normal genotype) will either be carriers or homozygous for the normal allele (Fig. 5). If a man happens to have an X-linked recessive disorder, he cannot pass it down to his sons, whom he must pass his Y chromosome down to. Therefore all his sons will be unaffected, but his daughters may be carriers.
The above pedigree may seem very complex, but we can break it down to understand some basic principles. Firstly, all affected individuals are males and they are inheriting this disorder from parents, both of which are not affected. If this disorder had an autosomal recessive inheritance, it would be seen in both male and female descendants. Because it is exclusively seen in males, we can safely presume the disorder is X-linked recessive.
4. X-linked Dominant
Most X-linked disorders are recessive, but a few are dominant. This means that the parent who has the trait also has the disorder, and when they pass this trait down the children who receive it will be affected as well (Fig. 6).
A woman with an X-linked dominant disorder passes it down to her sons and daughters equally. One of the biggest hints suggesting X-linked dominant disorders is that a man who has an X-linked dominant disorder must pass it down to all his daughters, as that is the only chromosome he can give them.
5. Y-linked Disorders
Very few disorders or traits have been discovered to be Y-linked. In fact, the preponderance of disorders that primarily affect men is typically due to the presence of a single X-chromosome, such that whatever disordered trait is on that chromosome cannot be masked by the normal trait that would be on a paired X-chromosome in females.
Ultimately, we can know Y-linked traits because they never occur in females, only in males (Fig. 7). And an affected male must pass the trait down to all his sons. Some forms of deafness are Y-linked.
6. Mitochondrial Inheritance
Mitochondrial inheritance is maternal, meaning we get our mitochondria from our mothers. Thus, an affected woman passes down a trait to all her children, and only her daughters can pass it on to their children (Fig. 8).
Human Pedigree Analysis: Problem Sheet
Now that we know the six major groupings of pedigree analysis, we can develop a problem sheet - in the form of a table- to help us consolidate the principles of each pedigree (Table 1).
| Autosomal recessive | |
| Autosomal dominant | |
| X-linked recessive | |
| X- linked dominant | |
| Y- linked | |
| Mitochondrial inheritance |
Table 1: Hints for pedigree analysis problem sheets. Chisom, Vaia.
Pedigree Analysis - Key takeaways
- Pedigrees can help us to analyze the inheritance patterns of many traits
- Pedigrees are typically used in the setting of genetic disorders
- The most common inheritance patterns include autosomal recessive , autosomal dominant and X-linked recessive .
- Some other less common inheritance patterns include X-linked dominant , Y-linked and mitochondrial inheritance .
- To solve a pedigree analysis , first look for dominance, than look for possible sex-linkage.
Flashcards in Pedigree Analysis 50
What kind of inheritance pattern does sickle cell anemia have?
autosomal recessive
Which of these inheritance patterns is the rarest for genetic disorders?
What is the name of the disorder that causes severe respiratory illnesses, is very common in North European individuals, and is due to a defect in chloride channels?
Cystic Fibrosis
What two tools are used frequently to test for the presence of trisomy in a fetus while they're still in utero?
Chorionic Villus Sampling and Amniocentesis
What is the name of the tool/method that allows us to visualize chromosomes to examine their size, number, and shape?
Karyotyping
What is the inheritance pattern of Hemophilia?
x-linked recessive

Learn with 50 Pedigree Analysis flashcards in the free Vaia app
We have 14,000 flashcards about Dynamic Landscapes.
Already have an account? Log in
Frequently Asked Questions about Pedigree Analysis
how to solve pedigree analysis
To solve a pedigree analysis, we must first determine if the trait is dominant or recessive. Look at parents and children's state to determine this.
what is the importance of pedigree analysis
Pedigree analysis is important because it helps us to predict the likelihood of future offspring having a disorder.
what is pedigree analysis
A pedigree analysis is a visual depiction of the genetic states of members of a family - carriers, affected, or completely unaffected.
How do you analyze a pedigree
Analyze a pedigree by first determining the dominance of a trait, and then determining its sex-linkage.
Test your knowledge with multiple choice flashcards

Join the Vaia App and learn efficiently with millions of flashcards and more!
Keep learning, you are doing great.
Discover learning materials with the free Vaia app

Vaia is a globally recognized educational technology company, offering a holistic learning platform designed for students of all ages and educational levels. Our platform provides learning support for a wide range of subjects, including STEM, Social Sciences, and Languages and also helps students to successfully master various tests and exams worldwide, such as GCSE, A Level, SAT, ACT, Abitur, and more. We offer an extensive library of learning materials, including interactive flashcards, comprehensive textbook solutions, and detailed explanations. The cutting-edge technology and tools we provide help students create their own learning materials. StudySmarter’s content is not only expert-verified but also regularly updated to ensure accuracy and relevance.
Vaia Editorial Team
Team Pedigree Analysis Teachers
- 11 minutes reading time
- Checked by Vaia Editorial Team
Study anywhere. Anytime.Across all devices.
Create a free account to save this explanation..
Save explanations to your personalised space and access them anytime, anywhere!
By signing up, you agree to the Terms and Conditions and the Privacy Policy of Vaia.
Sign up to highlight and take notes. It’s 100% free.
Join over 22 million students in learning with our Vaia App
The first learning app that truly has everything you need to ace your exams in one place
- Flashcards & Quizzes
- AI Study Assistant
- Study Planner
- Smart Note-Taking

Privacy Overview

An official website of the United States government
The .gov means it’s official. Federal government websites often end in .gov or .mil. Before sharing sensitive information, make sure you’re on a federal government site.
The site is secure. The https:// ensures that you are connecting to the official website and that any information you provide is encrypted and transmitted securely.
- Publications
- Account settings
Preview improvements coming to the PMC website in October 2024. Learn More or Try it out now .
- Advanced Search
- Journal List
- CBE Life Sci Educ
- v.21(1); Spring 2022
Secondary Students’ Reasoning on Pedigree Problems
Justin timm.
a Biology Education Research and Learning Lab, University of Duisburg-Essen, 45141 Essen, Germany
Katharina Wools
Philipp schmiemann, associated data.
Pedigree problems are typical genetics tasks in schools. They are well suited to help students learn scientific reasoning, representing realistic genetic problems. However, pedigree problems also pose complex requirements, especially for secondary students. They require a suitable solution strategy and technical knowledge. In this study, we examined the approaches used by N = 89 secondary school students when solving two different pedigree problems. In our qualitative analysis of student responses, we examined how two groups of secondary students with varying degrees of experience in genetics constructed arguments to support their decisions. To do so, we categorized I = 516 propositions from students’ responses using theory- and data-driven codes. Comparison between groups revealed that “advanced genetics” students ( n = 44) formulated more arguments, referred more frequently to specific family constellations, and considered superficial pedigree features less often. Conversely, “beginning genetics” students did not use a conclusive approach of step-by-step falsification but argued for the mode of inheritance they believed was correct. Advanced genetics students, in contrast to beginners, to some extent used a falsification strategy. Finally, we demonstrate which family members students used in their decisions and discuss a variety of typical but unreliable arguments.
INTRODUCTION
Pedigrees or family trees are a highly standardized graphical representation of family medical histories ( Bennett et al. , 1993 , 1995 , 2008 ; Bennett, 2010 ). They are used in genetic counseling and research to determine genetic influence on a trait, to identify patterns of inheritance, to calculate the disease risk for an individual, and for multiple other reasons ( Bennett, 2010 , presents an extensive list). Moreover, “human pedigree analysis is a typical example of genetics problem solving” ( Corbett et al. , 2010 , p. 221). In this paper, we analyze secondary students’ argumentation and strategic approaches when solving pedigree problems, as these are well suited for learning scientific reasoning. We used a qualitative approach and compared two groups of secondary students with divergent levels of genetics education.
Pedigree Problems
Pedigree problems are frequently used in genetics education. A typical pedigree problem consists of a representation of a human genetic pedigree with a task aim set to identify the mode of inheritance that is (most likely) represented. Typically, four modes of inheritance are differentiated: autosomal dominant inheritance, autosomal recessive inheritance, X-linked dominant inheritance, and X-linked recessive inheritance.
In autosomal dominant inheritance, the presence of a single dominant allele results in the expression of the trait for which the allele codes. Each person with the trait has at least one affected parent. In autosomal recessive inheritance, the trait is expressed only if a person has inherited one allele from each parent. Heterozygous individuals do not develop the trait. They can, however, still inherit the affected allele and are, therefore, called carriers. An X-linked dominant trait is encoded by an allele located on the X chromosome. Males have only one X chromosome. As a result, all their daughters but none of their sons will inherit the trait. For females, however, both sons and daughters can inherit the trait. In X-linked recessive inheritance, only the absence of an unaffected allele results in the expression of the trait. Males are more likely to be trait carriers, because they cannot compensate for an inherited allele.
A complete solution to a pedigree problem requires the identification of the underlying mode of inheritance; it consists of a decision in favor of one pattern and, in the best case, evidence for the exclusion of all alternative patterns. Therefore, an exhaustive solution requires testing of multiple hypotheses ( Hackling and Lawrence, 1988 ; Smith and Good, 1984 ; Hackling, 1994 ). In a systematic approach, all modes of inheritance are tested to determine whether they can explain the trait’s distribution. Basically, this process constitutes a search for inconsistencies in which family constellations are identified as cues that would not be possible assuming one of the modes of inheritance in question; in this context, we define the genetic family constellation as the elementary informative unit consisting of father, mother, and one child. An inconsistency such as an affected child with unaffected parents, for example, proves that the trait is not dominant. Smith (1988) calls the combination of such an informative family constellation and the conclusion to be drawn from it a production rule. These production rules consist of condition–action pairs for which identifying a condition—for example, the occurrence of unaffected parents with an affected child—leads to a specific action, in this case, the conclusion that the trait cannot be dominant ( Smith, 1988 ). In the case of a pedigree problem with a clear solution, all but one mode of inheritance can be excluded by using a combination of several of these rules or by systematically assigning genotypes to all individuals and looking for contradictions. However, pedigree problems seem to be challenging for students ( Hackling and Lawrence, 1988 ; Smith, 1988 ; Hackling, 1994 ; Knippels et al. , 2005 ). Various difficulties have been described. In the study by Smith (1988) , unsuccessful participants tended to consider the first plausible option as an acceptable answer and used an incomplete falsification strategy during pedigree analysis. They rarely tested hypotheses using genotypes and often used imprecise or incorrect logic. They tended to identify a larger number of noncritical cues; they then based decisions on these cues and tended to be unable to make use of production rules. In short, they failed because their rules were incomplete or incorrect, inappropriately applied, or did not lead to any conclusions. In addition, they based decisions on genetic ratios, which is usually not reasonable, as pedigrees are normally too small to comply with statistical distributions. Hackling and Lawrence (1988) demonstrated that university genetics professors’ solutions to pedigree problems were more complete than those of both novice and competent human biology university students. The experts (professors) identified a larger number of critical cues and varied their hypothesis-testing strategy according to the prevalence of the trait. They were likely to use genotypes to test hypotheses for a common trait, but relied more on critical cues for rare traits. In contrast, in the case of students, this difference was not noticeable. In another study, Hackling (1994) found that, in a group of novice students, almost no one succeeded in correctly interpreting cues with respect to X-linked inheritance. In addition, a significant proportion of this group was unable to assign genotypes to a pedigree that showed the occurrence of an X-chromosomal dominant trait. In a study by Corbett et al. (2010) , if-then statements (which they call “cognitive statements”) to determine carrier status in case of an X-linked recessive inheritance caused the greatest difficulty for university students.
Pedigree Problems as One Type of Genetic Problem
Pedigree problems are just one type of genetic problem that requires problem-solving skills. They are classified as rather difficult problems, because they require a fairly challenging “effect-to-cause” reasoning between generations, whereas the simplest problem type, in contrast, only requires “cause-to-effect” reasoning within generations ( Stewart, 1988 ; Hickey et al. , 2000 ; Tsui and Treagust, 2010 ). Pedigree problems belong to the area of Mendelian genetics, which is based on the principles of meiosis ( Stewart et al. , 2005 ). Collins and Stewart (1989) identified four typical classes of problems within the area of Mendelian genetics, which they grouped into inheritance pattern problems and modifier problems: Inheritance patterns describe the relationship between genotype and phenotype. Typical patterns are simple dominance, codominance, and multiple alleles (which is a combination of the previous two; Collins and Stewart, 1989 ). Modifiers affect the inheritance pattern by altering or constraining the way alleles are transmitted. They can be present or absent but are bound to an inheritance pattern; they cannot exist without it. Possible modifiers include X-linkage, genetic linkage (when genes for two traits are closely to each other located on a chromosome), and lethal alleles ( Collins and Stewart, 1989 ). According to Collins and Stewart (1989) , the inheritance patterns of simple dominance, codominance, and multiple alleles and X-linkage as a modifier are all typically considered in introductory courses.
In pedigree problems, however, simple dominance and X-linkage are predominantly considered. While in other tasks the modifier “X-linkage” may be present or absent, this does not apply to pedigree problems: Once introduced, all four modes of inheritance (autosomal dominant inheritance, autosomal recessive inheritance, X-chromosomal dominant inheritance, and X-chromosomal recessive inheritance) are almost always considered. As a result, a complete solution should typically take all four modes of inheritance into account. In any case, pedigree problems are well suited for inquiry-based genetics education ( Stewart et al. , 2005 ).
Argumentation and Content Knowledge
Tasks involving pedigrees require problem-solving skills, because students need to use content knowledge, formulate hypotheses, and apply one or multiple solution strategies ( Aznar and Orcajo, 2005 ). They need to generate conclusive arguments to prove their problem solutions, and argument quality is presumed to be related to content knowledge ( Sadler and Zeidler, 2005 ; Sadler and Fowler, 2006 ; McNeill et al. , 2006 ; Aufschnaiter et al. , 2008). But what makes a conclusive argument? A simple argument consists of a claim supported by data as evidence ( Toulmin, 2003 ). However, more complex arguments can have a number of other components. Following the structure of Toulmin (2003) , arguments can also include warrants and backings, qualifiers, and rebuttals. Warrants are generally applicable rules and laws that may be included to legitimize the step from data to claim. Backings can be cited to legitimize the applicability of the warrants. As a simplification of the Toulmin model, warrants and backings can be summarized under the term “reasoning” ( McNeill et al. , 2006 ; Berland and McNeill, 2010 ); that is, reasoning encompasses justifying how the evidence chosen leads to the claim made. The model of Toulmin (2003) also contains qualifiers and rebuttals: Qualifiers modify the strength of a claim by taking into account rebuttal(s) in the form of exceptions and conditions under which the warrant does not apply. Transferred to our context, a good and comprehensive argument against a particular inheritance might look like this: The trait at hand cannot be inherited in an autosomal dominant manner (claim), because a couple without the trait has an affected child (evidence). This rules out autosomal dominant inheritance, because at least one parent would also have to be an allele carrier for the child to inherit a dominant allele. However, this parent would also have to be affected, because this dominant allele would have led to the expression of the corresponding phenotype (warrant or reasoning). As a result, to formulate a complete explanation to a pedigree problem, students must formulate conclusive arguments against three of the four relevant modes of inheritance based on content knowledge and evidence available in the pedigree.
Regardless of content knowledge, students struggle to formulate coherent arguments. They seem to have problems citing sufficient evidence ( Sandoval and Millwood, 2005 ) and justifying why evidence leads to specific claims ( McNeill et al. , 2006 ). According to three different learning progressions ( Songer et al. , 2009 ; Berland and McNeill, 2010 ; Lee et al. , 2014 ), students’ scientific argumentations and explanations can be categorized according to their complexity: Basal scientific argumentations and explanations are based on unsubstantiated claims, while more elaborated explanations and argumentations are characterized by claims that are justified by evidence. Arguments that entail warrants and backings (reasoning) on how evidence and assertion are related indicate an even more complete or complex level of scientific argumentation or explanation.
According to the model of Shea et al. (2015) , the interplay of content knowledge, argumentation skills, and situational features accounts for genetics literacy. Therefore, situational features may also influence genetic reasoning and thus problem solving ( Freidenreich et al. , 2011 ; Shea et al. , 2015 ; Schmiemann et al. , 2017 ).
To clearly determine the mode of inheritance in the case of a pedigree problem, all alternative modes of inheritance must be excluded with justification. The literature suggests that even university students often struggle with pedigree problems and fail to form conclusive and complete solutions. Nevertheless, students are already expected to solve pedigree problems in secondary school. With this study, we wanted to explore secondary school students’ approaches to solving pedigree problems and how they constructed arguments to support their decisions. Our research questions were: What arguments do secondary school students generate when analyzing human genetic pedigrees and what evidence are the students’ arguments based on? Do their arguments differ depending on their level of genetics education?
METHODOLOGY
Instrument and data collection.
In pursuit of our aim, we used paper-and-pencil tests with pedigree problems as open-ended items ( Wools, 2018 ). In these tests, students were asked to identify the present mode of inheritance and to write down their problem-solving approach as coherently as possible for each pedigree problem. At the end of each pedigree problem, students had to mark the correct mode of inheritance. To visualize these problems, we used two pedigrees ( Figure 1 ), which were generated semi-automatically based on matrices ( Surmann, 2017 ) and plotted with the R package kinship2 ( Therneau and Sinnwell, 2015 ). We selected autosomal pedigrees in which the pattern of inheritance can be determined without any doubt, as other pedigree problems may be too complex for students with little experience in genetics. For precise communication, each person in the pedigree was labeled with a unique identification number, and students were requested to always indicate these numbers when referring to specific family constellations. In the paper-and-pencil tests, each person in the pedigree was also given a name to prevent subjects from confusing the symbols for male and female; this additional information, however, has been omitted here to keep the pedigrees minimalistic.

Pedigrees used in the paper-and-pencil tests.
The trait mapped in the first pedigree ( Figure 1a ) is inherited in an autosomal recessive pattern. The mode of inheritance can be determined as autosomal recessive based on a single family constellation. Unaffected parents (no. 107 and no. 108) have an affected daughter (no. 113). Therefore, the trait can neither be autosomal dominant nor X-linked dominant, because at least one parent would have to be affected too. In addition, inheritance cannot be X-linked recessive, as the daughter would have inherited an allele from each parent. However, in this case, the father would have been affected too, because men are hemizygous (have only one allele). The pedigree shows inheritance across three generations and includes 14 persons in total, with three females and one male being affected.
The second pedigree problem involves an autosomal dominantly inherited trait ( Figure 1b ). This pedigree comprises six persons spread over three generations; three of the individuals (two males and one female) are affected. In this case, two family constellations are needed to determine that the inheritance is autosomal dominant. That two affected parents have an unaffected son rules out both recessive modes of inheritance, because parents no. 103 and no. 104 could not have possessed or transmitted any allele that is not associated with the trait, and so their son (no. 111) should have been affected as well. At the same time, an X-linked dominant inheritance is ruled out by the constellation of the unaffected daughter (no. 105) and the affected father (no. 101): Because men have only one X chromosome, the daughter would have inherited the relevant allele in any case and should, therefore, have been affected too.
For this study, we analyzed paper-and-pencil tests of 89 secondary students. In German secondary schools, pedigree problems are generally part of a basic genetics unit as well as an advanced genetics unit. The advanced unit takes place at the upper secondary level, usually 2 years after the basic unit. Although pedigree problems are thus part of both units, they are treated in much more detail in the advanced genetics unit. To cover a maximum of the variance expected in secondary school, we decided to include students who have only basic experience with pedigree problems as well as students with advanced experience. We examined 45 secondary students (M Age = 14.9), who completed a basic genetics unit, and 44 secondary students (M Age = 16.8) who had participated in the advanced genetics unit. Although we do not possess detailed information about the scope and process of the genetics units in the participating classes, pedigree problems are typical tasks covering scientific inquiry in genetics. For instance, pedigree analysis can be covered by an exemplary task within the educational standards for middle school graduation, which define an expected average achievement level for students at the end of lower secondary education ( Standing Conference of the Ministers of Education and Cultural Affairs of the Länder in the Federal Republic of Germany, 2005 ). In that task, students are expected to be able to determine the mode of inheritance in the presented pedigree by, among other things, ruling out an X-linked inheritance. Furthermore, pedigree problems are a typical task for the biology exams in the Abitur, the final examination at the end of the higher secondary level. According to the curriculum, upper secondary students are expected to formulate hypotheses on X-linked and autosomal modes of inheritance and justify the hypotheses with available data based on meiosis ( Ministry for School and Further Education of the State of North Rhine-Westphalia, 2014 , p. 32). Students taking an advanced course (in contrast to those who take a basic course) should additionally be able to consider two-factor analysis, genetic linkage, and crossing-over thereby. As a result, students from both groups should be familiar with all four modes of inheritance and with pedigree analysis in principle. Nevertheless, it is reasonable to assume that the students who participated in the advanced course have more differentiated knowledge and more procedural skills, because they had already attended the basic genetics unit a few years earlier. The advanced course can therefore build on these basics. Following ethical and legal guidelines, all participants were informed about the aim and procedure of the study in advance and participated voluntarily. The results were not used for evaluation or grading. Only necessary data were collected, and they were treated in accordance with the local data-protection laws. As the study was conducted in Germany, students answered in German; quotations in this article were translated into English by the authors.
Coding and Analysis
To answer our research question, we performed a content analysis of students’ reasoning on pedigree problems using MAXQDA 2018 ( VERBI Software, 2018 ). For this purpose, the students’ written answers for the open-ended tasks were transcribed. We defined propositions as coding units to analyze students’ reasoning at the level of individual conclusions ( Hackling and Lawrence, 1988 ). We developed a codebook based on theory and data as a guide for analyzing the students’ answers. Therefore, the codebook ( Table 1 ) contained theory-driven and data-driven codes ( DeCuir-Gunby et al. , 2011 ), which we present in detail here.
Variables and categories used to characterize students’ reasoning on pedigree problems (a complete version including code descriptions and examples can be found in the Supplemental Table 1)
| Variable | Categories |
|---|---|
| Type of statement | Description |
| Allegation | |
| Clue | |
| Proof | |
| Other | |
| Pedigree feature | Phenotypic family constellation |
| Genotypic family constellation | |
| Proportion of affected persons | |
| Gender ratio among affected | |
| Distribution of affected over generations | |
| Pedigree size or structure | |
| Other | |
| Missing | |
| Claim | Autosomal dominant |
| Autosomal recessive | |
| X-linked dominant | |
| X-linked recessive | |
| Autosomal | |
| X-linked | |
| Y-linked | |
| Dominant | |
| Recessive | |
| Other | |
| Orientation | Confirmation |
| Refutation | |
| Conclusiveness | Conclusive |
| Indeterminable | |
| Inconclusive |
First, we categorized whether the proposition was at all a conclusion or what type the proposition was. In this regard, we distinguished five types of propositions—“proof,” “clues,” “allegations,” “descriptions,” “other”—based on the present transcripts but strongly aligned with the theoretical frameworks of Toulmin (2003) and Smith (1988) . Based on the classification of Toulmin (2003) , a full argument contains at least a claim based on data. In pedigree analysis, typical arguments formulate claims regarding the confirmation or exclusion of at least one type or mode of inheritance based on data from the pedigree as evidence; for example, “the mode of inheritance is autosomal [claim], as both sexes are affected [data]” (Student 1009|Line 40). Therefore, arguments regarding the mode of inheritance represent condition–action pairs, as described by Smith (1988) . However, these arguments can also be further classified from a scientific perspective. Arguments regarding the mode of inheritance can represent either clear-cut decisions or indications for or against individual modes of inheritance. For example, in the case of X-linked recessive inheritance, more males tend to be affected than females. If such a distribution is present, this can therefore be an indication of an X-linked recessive inheritance, but it is not suitable as definitive proof. To account for this distinction, we have used two categories: proof and clues. In proof-type arguments, one mode of inheritance is unambiguously confirmed or ruled out, whereas arguments that provide evidence for or against modes of inheritance without making a clear decision were called clues. Although technical in nature, this distinction can also be understood in terms of Toulmin’s (2003) structure. Arguments that provide evidence for or against single heritages but do not allow a clear decision usually contain what Toulmin (2003) calls a qualifier. This is an optional argument component that indicates some uncertainty, as we can see in the following statement: “Again, both sexes are affected [data], which again indicates [qualifier] autosomal inheritance [claim]” (1012|41). Beyond these two central categories, three other types of statements were distinguished that do not constitute complete arguments according to Toulmin (2003) . In this regard, two categories—“allegations” and “descriptions”—represent incomplete arguments, because they consist of only one element of Toulmin’s (2003) classification each. Allegations represent stand-alone claims that do not contain any data as evidence. Descriptions mention data from the pedigree but do not contain a claim. Finally, statements that did not fit into one of the first four categories were classified as “other.”
To differentiate the various types of pedigree features students cited as data in their arguments (or descriptions), we distinguish eight categories based on students’ statements and results from previous research. A typical approach during pedigree analysis is to test multiple hypotheses based on informative family constellations or by assigning genotypes ( Hackling, 1994 ). Based on the written arguments, we cannot determine whether students identified a family constellation by recalling memorized production rules or assigning genotypes. Nevertheless, we were able to distinguish whether they argued using genotypes or phenotypes. In cases in which students mentioned a particular family constellation in a statement, we classified the statements according to whether the students were referring to the phenotypic level (coded as “phenotypic family constellation”) or genotypic level (coded as “genotypic family constellation”). Another category we found in our data regarding pedigree analysis includes statements referring to the “proportion of affected persons” in relation to the total number of persons. This category addresses the misconception that dominance and allele or trait frequency are linked, which has been described in previous work, not just in the context of pedigree analysis (e.g., Smith and Good, 1984 ; Smith, 1988 ; Abraham et al. , 2014 ). This becomes evident in the following example: “Since very few people are affected, the inheritance is recessive” (1003|34). The starting point for another pair of categories was a case study presented by Hackling (1994) , in which one subject determined the mode of inheritance to be autosomal dominant because there were affected individuals in each generation (mode of inheritance cannot be recessive, must be dominant) and males and females were affected (mode of inheritance cannot be X-linked). Based on the intersection of Hackling’s (1994) descriptions and our data, we created categories to capture the cases in which students quote the “gender ratio among affected” individuals or the “distribution of the affected across generations.” Not captured by the categories mentioned so far is the fact that students also referred to basic features of the pedigree, such as the number of generations represented. Therefore, we added a category to cover statements in which students referred to the overall “pedigree size or structure.” In addition, sometimes a datum from the pedigree was “missing” or students mentioned “other” pedigree features we were unable to categorize more accurately.
To analyze students’ claims more precisely, we introduced categories based on content knowledge and added all relevant modes of inheritance (“autosomal dominant,” “autosomal recessive,” “X-linked dominant,” “X-linked recessive,” or “Y-linked”) as well as more general conclusions on heredity (“dominant,” “recessive,” “autosomal,” or “X-linked”) as categories. Here too, claims that could not be assigned to any of these categories were coded as “other.” While the previous two variables account for the biology-specific content of the propositions, we also analyzed students’ strategic procedures. The extent to which the students used a falsification strategy ( Smith, 1988 ) could be deduced by the orientation of their arguments. In this regard, we distinguished statements that entailed support for or confirmation of claims from those weakening or rejecting claims. As a result, we categorized each argument as “confirmation” or “refutation” with regard to the subject of the claim. The goal here was to investigate the extent to which students used a falsification strategy in their pedigree analyses. Finally, we classified each argument in terms of its conclusiveness, as either “conclusive” or “inconclusive” or, if necessary, as “indeterminable” based on expert knowledge.
To ensure the consistency of the coding, we selected a random set of 26.7% of all propositions, which were coded by two independent raters (JT and MG, a student assistent; O’Connor and Joffe, 2020 ). We calculated Cohen’s kappa ( Cohen, 1960 ) for all main categories of interest ( Table 1 ) and interpreted the values in the context of the limits described by Landis and Koch (1977) . We interpreted these values to check for inconsistencies and revised the coding manual accordingly ( O’Connor and Joffe, 2020 ). Then, the coding process was repeated until satisfactory agreement was obtained for all categories. According to the thresholds described by Landis and Koch (1977) , we achieved almost perfect agreement (0.8 < κ ≤ 1) for the pedigree feature (κ = 0.81), claim (κ = 0.92), and argument orientation (κ = 0.98). Substantial agreement (0.6 < κ ≤ 0.8) was achieved for type of statement (κ = 0.75) and argument conclusiveness (κ = 0.77). We considered these values to be satisfactory, and so one rater (MG) coded all remaining propositions to obtain the full set of data used in this study.
To analyze the coding results, we created tables to examine what kinds of propositions and which claims the students expressed for each pedigree. To examine the connections between students’ claims and the associated data, we created a mosaic plot. Mosaic plots are used to visualize contingency tables and consist of tiles that represent the individual frequencies through tile size ( Hofmann, 2008 ).
In addition, we determined which pedigree members students mentioned in their reasoning by calculating frequencies for all pedigree members’ identification numbers. To visualize these data, we drew the individual frequencies directly into the pedigrees. To obtain comprehensible diagrams, we applied the principles of heat maps and bubble charts to present our data. This means that the relative frequencies are represented by the size and color of the bubble drawn on top of each family member’s symbol. A large red bubble indicates that a person has been mentioned frequently, while a small yellow bubble indicates that the person has been mentioned very rarely.
The total data set consists of 516 propositions, on average 6.1 per student (excluding five students who did not answer at all). Overall, the number of propositions made was slightly higher for the first task (the autosomal recessive pedigree) than for the second task (the autosomal dominant pedigree). The proportion of statements representing full arguments (clues or proof) was considerably higher in the group of advanced genetics students, χ 2 (1) = 36.68, p < 0.001 (see Table 2 for detailed results).
Number and types of propositions for both groups of students a
| Type of proposition | ||||||||
|---|---|---|---|---|---|---|---|---|
| Group | Subjects | Task/pedigree | Propositions | Propositions PP | Descriptions | Allegations | Arguments | Other |
| Beginning genetics students | 39 | Autosomal recessive inheritance | 134 | 3.44 | 64(47.8%) | 14(10.4%) | 48(35.8%) | 8(6%) |
| 40 | Autosomal dominant inheritance | 113 | 2.83 | 46(40.7%) | 16(14.2%) | 44(38.9%) | 7(6.2%) | |
| Advanced genetics students | 43 | Autosomal recessive inheritance | 153 | 3.56 | 34(22.2%) | 9(5.9%) | 93(60.8%) | 17(11.1%) |
| 43 | Autosomal dominant inheritance | 116 | 2.70 | 18(15.5%) | 8(6.9%) | 80(69%) | 10(8.6%) | |
a Students who did not respond to a task were excluded from the individual calculation. Propositions PP: Propositions per person.
Beginning Genetics Students Do Not Use a Falsification Strategy
To examine whether students used a falsification strategy to identify the mode of inheritance by ruling out all alternative modes, we analyzed their arguments in multiple regards. Beginning genetics students predominantly (92.6%) formulated confirmatory statements; for example: “Both men (103) and women (104) have the trait, so it is autosomal” (1023|41). Advanced genetics students, in contrast, formulated confirming (58.9%) as well as falsifying arguments (41.1%); for example: “Gonosomal dominant can be excluded on individuals 101, 102, and 104” (B15|41). This difference is significant, χ 2 (1) = 40.24, p < 0.001, and demonstrates that the advanced genetics students resorted to a falsification strategy at least temporarily, while the beginning genetics students made a straightforward decision on the mode of inheritance. However, these two quotes also show a difference in scale. Arguments that refer to a specific mode of inheritance—for example, autosomal dominant or X-linked recessive inheritance—were rare in the group of beginning genetics students but rather common in the group of advanced genetics students (see Table 3 ). In both groups, autosomal recessive inheritance was the most frequently mentioned specific mode of inheritance. In addition, arguments that either indicated whether inheritance was sex-linked or drew a conclusion about the trait’s dominance without naming a specific inheritance mode were common in both groups (see Table 3 ). To examine the quality of an argument, however, we found it necessary to look at the data used to substantiate the claims.
Subjects students refer to in their conclusions by task and group a
| The claim refers to … | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Group | Subjects | Task/pedigree | Propositions | Dominant inheritance | Sex-linked Inheritance | AD | AR | XD | XR | Y | Other |
| Beginning genetics students | 23 | Autosomal recessive inheritance | 48 | 26(54.2%) | 10(20.8%) | 0(0%) | 2(4.2%) | 1(2.1%) | 2(4.2%) | 0(0%) | 7(14.6%) |
| 26 | Autosomal dominant inheritance | 44 | 23(52.3%) | 12(27.3%) | 1(2.3%) | 4(9.1%) | 0(0%) | 0(0%) | 0(0%) | 4(9.1%) | |
| Advanced genetics students | 39 | Autosomal recessive inheritance | 93 | 34(36.6%) | 16(17.2%) | 5(5.4%) | 11(11.8%) | 6(6.5%) | 9(9.7%) | 1(1.1%) | 11(11.8%) |
| 37 | Autosomal dominant inheritance | 80 | 31(38.8%) | 16(20%) | 7(8.8%) | 6(7.5%) | 9(11.2%) | 4(5%) | 0(0%) | 7(8.8%) | |
a Students who did not respond to a task or did not make any analytical statements there were excluded from the individual calculation. AD, autosomal dominant inheritance; AR, autosomal recessive inheritance; XD, X-linked dominant inheritance; XR, X-linked recessive inheritance; Y, Y-linked inheritance.
A Variety of Decisions Are Based on Superficial Pedigree Features
Students referenced a variety of pedigree features as data to prove their claims; for example, the gender ratio among affected persons, the proportion of affected persons, or the distribution of the trait over generations. We call these features “superficial,” because they are easily noticed but allow no or only vague conclusions (clues). Family constellations, either at the phenotypic or genotypic level, form the counterpart. They must be remembered or analyzed precisely. Unlike “superficial” features, family constellations allow clear decisions. In fact, many students described specific family constellations, often at the phenotypic level, sometimes at the genotypic level. In total, 89.9% of all students referred at least once to a specific family constellation. However, not every proposition contains a conclusion. To identify which pedigree features were mentioned in connection with which type of inheritance, we analyzed the relationship between claim (on the mode of inheritance) and data mentioned (the pedigree feature to justify that claim; Figure 2 ).

Mosaic plot showing the structure of students’ arguments. The relationship between the inheritance mentioned in the argument and the data used to prove that claim is shown separately for each group. The diagram maps all possible combinations of pedigree features and claims regarding the inheritance, while the area of each box represents how often that specific combination was found among students’ statements. Thin lines without percentages indicate that a specific combination was not found. AD, autosomal dominant inheritance; AR, autosomal recessive inheritance; XD, X-linked dominant inheritance; XR, X-linked recessive inheritance; Y, Y-linked inheritance.
It becomes apparent through analysis of the mosaic plots ( Figure 2 ) that advanced genetics students based their decisions regarding the mode of inheritance primarily on specific family constellations within the pedigree. The proportion of statements referring to family constellations was significantly larger in the group of advanced genetics students (75.5%) compared with the group of beginning genetics students (46.9%), χ 2 (1) = 18.02, p < 0.001. In addition, a significantly larger proportion of advanced genetics students’ arguments focused on specific modes of inheritance (37.4% compared with 12.3% of basic genetics students’ arguments), χ 2 (1) = 15.11, p < 0.001. The distribution of affected over generations and the gender ratio among affected individuals were cited fairly often, each in a specific context. The distribution of affected over generations was primarily used to prove the trait’s dominance (12 of 15 mentions were associated with this kind of claim), and the gender ratio among affected individuals was primarily mentioned in connection with sex linkage. Indeed, most arguments by beginning genetics students regarding sex linkage were based on the gender ratio among affected, while even references to specific family constellations were rare in this case. Compared with advanced genetics students, beginning genetics students referred significantly more often to the gender ratio among affected in their arguments on sex linkage, χ 2 (1) = 4.69, p = 0.030. Overall, beginning genetics students argued more frequently on the basis of superficial pedigree features, χ 2 (1) = 21.18, p < 0.001, and less frequently on the basis of genotypes, χ 2 (1) = 7.84, p = 0.005. A rare kind of data, primarily used by beginning genetics students (eight out of 10 mentions occurred in this group) was the proportion of those affected. It was cited primarily in relation to the trait’s dominance (seven of the total 10), which, however, is not appropriate, because trait prevalence depends on allelic frequency alone and does not allow any indications of dominance. Accordingly, arguments based on this kind of evidence are inconclusive in any case. The conclusiveness of arguments relating to specific family constellations, in contrast, cannot be assessed across the board. An extended mosaic plot showing which arguments were classified as correct and at what frequency can be found in the Supplemental Material. An argument does not need to be conclusive just because a particular constellation in the pedigree is pointed out. To analyze in detail which family members the students referred to, we used the identification numbers of the pedigree members. The frequency of the mentions was then drawn in the pedigree, separately for each task and student group ( Figure 3 ).

Representation of the frequency with which students mentioned individual family members in their arguments regarding the present mode of inheritance separately for each pedigree and group. The frequencies are shown in the pedigrees by the color and size of the points. Mentions of individual family members in the pedigree showing an autosomal recessive trait by (a) beginning genetics students and (b) advanced genetics students. Mentions of individual family members in the pedigree showing an autosomal dominant trait by (c) beginning genetics students and (d) advanced genetics students.
The first pedigree ( Figure 3a and b ) shows the inheritance of an autosomal recessive trait. In this case, all alternative modes of inheritance can be excluded by one family constellation (unaffected father no. 107 and mother no. 108 in combination with their affected daughter no. 113). In the group of beginning genetics students, however, the affected but uninformative person no. 104 was mentioned most often in arguments regarding inheritance (14 mentions; relative frequency: 100%). Also very frequently mentioned, in descending order, were the other affected no. 113 (93%), no. 105 (86%), no. 102, and no. 107 (each 64%). Reasonably often cited in arguments were person no. 108 (50%), the children of no. 103 (29%) and no. 104 (no. 110, no. 111, and no. 112; each 43%). In contrast, the picture is markedly different for advanced genetics students’ mentions. Most frequently mentioned was the informative person no. 113 (39 mentions; relative frequency: 100%) closely followed by the parents no. 107 (95%) and no. 108 (92%). The other affected family members, in contrast, are cited notably less frequently. Their mentions in descending order: no. 104 (56%), no. 102 (46%), and no. 105 (21%). Thus, the affected person no. 105 is mentioned less frequently than no. 101 (28%) and the children of no. 103 (also 21%) and no. 104 (no. 110: 26%; no. 111: 28%; no. 112: 23%). The relative frequency of mentions of the unaffected and uninformative individuals no. 106, no. 114, and no. 115 is low in both groups and does not exceed 15%. Overall, comparing the arguments referring to family constellations in terms of group membership reveals that the importance of the family constellation consisting of family members no. 107, no. 108, and no. 113 seems rarely to be recognized by the group of beginning genetics students. The advanced genetics students, in contrast, seem to recognize the importance of this family constellation.
The second pedigree ( Figure 3c and d ) shows the inheritance of an autosomal dominant trait. For the constellation given, the exclusion of all alternative modes of inheritance is possible based on two individual constellations. Based on the family constellation of the affected parents no. 103 and no. 104 in combination with their unaffected child no. 111, both recessive modes of inheritance are ruled out. An X-linked dominant inheritance can be excluded, because no. 105 is not affected but should have inherited the trait causing allele from father no. 101. Beginning genetics students refer in their arguments regarding the mode of inheritance mainly to the family constellation of mother no. 104 (26 mentions; relative frequency: 100%), father no. 103 (88%), and their son no. 111 (77%). Beyond that, only the affected male founder no. 101 is mentioned with noteworthy frequency (35%). The female founder no. 102 is cited rarely (12%), while the informative female no. 105 is mentioned only once (4%). Comparing these results with those of the advanced genetics students, it is noticeable that differences can be observed especially among those family members not mentioned by the beginning genetics students. In this group too, the informative family constellation of mother no. 104 (44 mentions; relative frequency 100%), father no. 103 (77%), and their son no. 111 (89%) is mentioned by far most frequently. However, the informative female no. 105 (36%) and her father no. 101 (45%) are also mentioned moderately often. In this group, least often mentioned is the female founder no. 102 (25%). These results indicate that both groups particularly recognize the family constellation of family members no. 103, no. 104, and no. 111 as relevant and formulate arguments based on this constellation. The second informative family constellation (no. 101, no. 102, and no. 105), necessary to rule out an X-linked dominant inheritance, is hardly ever mentioned by beginning genetics students. Advanced genetics students, in contrast, create arguments based on this family constellation too.
At the end of each task, students had to mark in a multiple-choice item which inheritance they thought was present. Overall, the beginning genetics students were less likely to answer correctly than the advanced genetics students. In the case of the autosomal recessive pedigree, the majority of students of both groups opted for the correct mode of inheritance (beginning genetics students: 65.9%; advanced genetics students: 88.1%). For the autosomal dominant pedigree, however, a majority of beginning genetics students again chose autosomal recessive inheritance (55.8%; only 32.6% decided correctly for autosomal dominant inheritance). Of the advanced genetics students, 56.1% correctly selected the autosomal dominant mode of inheritance. However, a correct decision in the multiple-choice item does not necessarily indicate that the decision is explained based on correct arguments. It can be stated that, in most cases, students chose the correct mode of inheritance if at least one of their arguments was found to be conclusive (see Figure 4 ). Even students who had not formulated any conclusive arguments in a pedigree problem sometimes identified the correct inheritance. The proportion is even quite large in the case of autosomal recessive inheritance, especially among advanced genetics students (see Figure 4 ). For an in-depth analysis of which arguments were rated as correct in which group, see Supplemental Figure 1.

Number of conclusive arguments per person and pedigree. Individuals were counted as a function of group membership. Colors indicate which proportion of individuals marked the correct inheritance in an attached multiple-choice item.
Students’ Use of Unreliable Arguments
As the previous results suggest, analytical statements made by advanced genetics students (60.7%) were more frequently conclusive compared with statements by beginning genetics students (29.3%), χ 2 (1) = 22.37, p < 0.001. In this section, we will analyze some of these inconclusive arguments in detail. We start by presenting some arguments in which students decide between dominant and recessive inheritance and then provide some arguments regarding sex linkage.
A typical conclusion based on the proportion of affected persons among all pedigree members was made by one student, who stated: “Since very few people are affected, the inheritance is recessive” (1003|34). In this statement, Student 1003 implies that autosomal dominant inheritance is indicated by the fact that a large number of people are affected. The proportion of people affected is highly dependent on chance and strongly influenced by the frequency of the trait-determining allele in the total population. Chance plays an important role, because pedigrees are usually too small for probability-based evidence. A carrier of an autosomal dominant trait may pass the trait to all his children or none; starting from one allele, all descendants could be affected or none of them. Nevertheless, the allelic frequency has an impact on how likely an autosomal recessive inheritance is ( Hackling and Lawrence, 1988 ). If the autosomal recessive trait is very common in the population, it is very likely that a considerable number of the family members who are marrying in are carriers. This would mean, in turn, that a large proportion of the pedigree members could be carriers. If an allele is very rare and many pedigree members are affected, an autosomal recessive inheritance is in fact rather unlikely, as it would require several unrelated persons marrying in to bear the rare allele. Because there was no information on allelic frequency for either pedigree problem here, the proportion of persons who are affected does not allow any conclusions to be drawn about the inheritance mode.
A typical conclusion based on trait distribution over generations is made by Subject B6: “First of all, it must be noted that the pedigree cannot be dominant, since the trait does not occur in every generation” (B6|40). What B6 observes here is the distribution of the trait over generations, which indeed can be used to rule out simple dominant inheritance (under the assumptions of full penetrance and no de novo mutations). A correct rule could state that a skipped generation rules out dominant inheritance, because that is about non-affected parents having an affected child. The comparison makes clear that the rule presented by Subject B6 deviates slightly from the appropriate rule and does not necessarily lead to correct results. For example, no statement can be made on the basis of a terminal generation without affected persons. While the slightly modified rule results in the correct decision for the autosomal recessive pedigree, Subject B6 de facto considers a dominant inheritance impossible for the autosomal dominant pedigree as well based on his rule. In return, his rule implies that a dominant trait is always transmitted. This is a recurring misconception, appearing, for example, in a statement by Subject B20: “The inheritance is also recessive, since none of the children [110, 111, 112] of 103 and 104 carry the trait. If the inheritance was dominant, 110, 111 or/and 112 would carry the trait, since the mother 104 carries the trait” (B20|41–42). In this statement, the student assumes that the trait cannot be dominant, because otherwise an affected mother would certainly have passed it on to at least one child. What we cannot say is which false assumption underlies this statement: On the one hand, the decision could be based on the expectation that a dominant trait will always necessarily be passed on to the next generation, which is not necessarily the case; on the other hand, the decision could be based on an incorrect understanding of probabilities, by assuming that, in the case of three children, at least one should be affected.
Based on the gender ratio, there were two different ways of reasoning evident. On the one hand, the distribution of the trait by gender was used as a qualitative measure: “The trait is inherited as an autosomal trait since both female and male persons are affected” (D11|40). In this argument, it seems irrelevant how many people of each gender are affected, as long as there is at least one each. On the other hand, the distribution of the trait depending on gender was used as a quantitative measure, too: “[T]he inheritance is gonosomal because almost only women are affected” (1003|36). In this case, an (extreme) imbalance was used as an argument for sex linkage.
In this study, we wanted to explore secondary school students’ approaches to solving pedigree problems and how they constructed arguments to support their decisions. For this purpose, we analyzed two pedigree problems representing autosomal modes of inheritance that should be known to all students. Overall, we identified a number of interesting findings.
While the students’ answers hardly differed in number of propositions by group, beginning genetics students generated significantly fewer arguments. A notable proportion of this group actually stated no arguments at all. In fact, many of the beginning genetics students’ statements were descriptive. Nevertheless, a considerable number of students correctly identified the autosomal recessive inheritance in the first pedigree, even in the absence of conclusive arguments (see Figure 4 ). There could be several reasons why the students’ answers turned out the way they did. First, some students may have found it difficult to write down their approaches. Among other things, a lack of expertise could have been responsible for this, because argumentation quality is presumed to be related to content knowledge ( Sadler and Zeidler, 2005 ; Sadler and Fowler, 2006 ; McNeill et al. , 2006 ; Aufschnaiter et al. , 2008). Second, some students may in fact have reached a conclusion on the mode of inheritance without citing any evidence or data. This would be in line with the proposed learning progressions on scientific argumentation, which assume that it is easier to formulate claims than to support them with evidence or data ( Songer et al. , 2009 ; Berland and McNeill, 2010 ; Lee et al. , 2014 ). In fact, this would also fit the observation that a substantial portion of students succeeded in correctly identifying the present mode of inheritance without having formulated any conclusive arguments. Third, some students may have considered a pedigree feature relevant but were unable to draw any conclusion from it. In this case, too, a lack of content knowledge could be the issue. Smith (1988) observed that individuals who were unsuccessful in pedigree analysis tended to quote affected and unaffected descendants of matings shown in the pedigree without drawing conclusions. He stated that “these individuals appear to be looking for recognizable patterns or other cues, but they either do not recognize significant cues or they do not know which of their patterns are meaningful” ( Smith, 1988 , pp. 416–417). Finally, some students could have assumed that a brief description of the pedigree was part of a conclusive pedigree analysis. However, beginning genetics students not only made more descriptive statements but also referred less frequently to specific family constellations and considered superficial pedigree features as data more often. For example, the numbers of affected and unaffected persons and the ratio between them were repeatedly mentioned, as was the number of generations. In particular, when deciding between autosomal or X-linked inheritance, beginning genetics students mainly relied on superficial features as evidence in their arguments. This is in line with the results of Hackling (1994) , who found that the majority of novice undergraduates lack the necessary content knowledge to reliably check for X-linked inheritance. Although the advanced genetics students referred to family constellations much more frequently, the only informative family constellation to rule out X-chromosomal dominant inheritance in pedigree 2 was almost never mentioned by beginning genetics students and only sometimes by advanced genetics students. In general, beginning genetics students focused primarily on affected family members whether or not they were informative; thus, a considerable proportion of students in this group did not seem to be able to distinguish between informative and uninformative family constellations. Advanced genetics students, in contrast, were particularly concerned with informative pedigree members, which indicates that they identified critical cues more often. This was true at least in the case of the autosomal recessive pedigree; for the autosomal dominant pedigree, however, the results were much less obvious. Therefore, a clear trend cannot be proven empirically or based on previous studies. Hackling and Lawrence (1988) observed that “experts recognized more critical cues than the students” (p. 537). In the study by Smith (1988) , however, unsuccessful and successful participants identified comparable numbers of critical cues, whereas unsuccessful participants identified noticeably more noncritical cues.
Nevertheless, identifying a critical cue does not necessarily mean drawing the right conclusions from it. This is independent of whether the cue was identified based on assigning genotypes or by recalling a memorized production rule. In both cases, genetic knowledge is required. When a certain condition occurs in the pedigree, for example, when two unaffected parents have an affected child, the corresponding action that a dominant inheritance is ruled out comes to pass. In our sample, only about every fourth argument by the beginning genetics students was technically sound, and even among advanced genetics students, more than one in three conclusions was incorrect. This observation is largely consistent with Smith’s (1988) comment that unsuccessful students often either do not use production rules at all or use them inaccurately, incompletely, or even incorrectly. Indeed, beginning genetics students often used superficial and uninformative pedigree features as data, which was probably the reason for the low number of valid conclusions.
Regarding the strategic approach, we highlight two points. First, advanced genetics students much more often used a falsifying procedure (cf. Hackling and Lawrence, 1988 ), whereas beginning genetics students almost exclusively generated confirmatory arguments. Second, but just as important, students in both groups primarily did not test the modes of inheritance individually, but rather decided between dominant and recessive, on the one hand, and autosomal and X-linkage, on the other hand.
Limitations
The analysis of pedigree problems is time-consuming, and all results described here were derived from the analysis of only one pedigree per type of autosomal inheritance. Based on the available data and results, it is therefore not possible to make any predictions about the general strategic approach of secondary students to other pedigrees, especially those that represent X-linked inheritance. Apparently, it is uncertain whether individual pedigree features influence students’ approaches and, if so, how. It is difficult to imagine varying multiple pedigree features systematically in order to investigate the effects of these variations on students’ procedure. The influence of features such as pedigree size or the proportion of affected can only be investigated, if at all, in large samples in which all subjects analyze only a subset of the pedigrees.
In our study, all students participated voluntarily, and their results were neither evaluated nor graded. Accordingly, it is reasonable to assume that some individuals may not have put full effort into analyzing both pedigrees and writing down their approaches. Nevertheless, we are convinced that we were able to uncover students’ typical patterns of reasoning and to identify what kind of evidence students typically use to justify their decisions. We examined a reasonably large sample of students in order to quantify our main results. However, this had an impact on how precisely we were able to analyze each student’s data. Because we analyzed written responses, it is likely that we missed some of the students’ thoughts. For example, we assume that they may not have written it down if they recognized a constellation as meaningful but were unable to draw any conclusions from it. To uncover such cases, one would have to use other methods that provide more insight into the thinking processes of the students, such as retrospective interviews, thinking aloud, or eye tracking ( Lai et al. , 2013 ). Because retrospective interviews and think-aloud protocols could be potentially biased by mental processes, eye-tracking data could be even better, because information intake is recorded directly and in an unfiltered manner. For this, however, the sample size would have had to be reduced significantly.
Implications for Teaching
Pedigrees are an excellent way to learn genetics, for multiple reasons, and are a core practice in genetic classes. Pedigree problems represent realistic scientific problems and illustrate real human genetic phenomena observable at the macroscopic level ( Tsui and Treagust, 2010 ). They are well suited for scientific reasoning and are appropriate to help students develop a comprehensive understanding of meiosis and Mendelian inheritance in humans.
Our findings offer some implications for what students should consider when analyzing pedigrees. First, a significant proportion of the participants’ conclusions were based on simplified rules, which may serve as a first indication but are not suitable for making clear decisions. We suppose these students were relying too much on their instincts. Students need to learn how to distinguish a presumption based on superficial features from a decision based on clear evidence. To achieve this, it is advisable to also examine pedigrees where the first impression is misleading; for example, a recessive pedigree in which most people are affected might be a good start. Second, we suppose that it is not necessary to start by memorizing all informative family constellations. Students should rather learn to analyze family constellations on demand, for instance, by assigning genotypes and using their knowledge of meiosis to test whether there is a contradiction for one mode of inheritance. Of course, this requires students to be able to correctly assign genotypes, even for sex-linked modes of inheritance. In a survey by Hackling (1994) , however, a large number of novice undergraduate university students were unable to assign genotypes for X-linked dominant inheritance accurately. We suppose that it would be worth considering practicing this method before considering X-linkage. Beginning genetics students made almost exclusively confirmatory statements, whereas a complete and conclusive procedure is usually characterized by the fact that all alternative inheritance modes are conclusively ruled out by testing multiple hypotheses ( Hackling and Lawrence, 1988 ). To encourage this approach, the task could be changed for practice, prompting students to explicitly exclude impossible inheritance modes one by one. Only when the procedure has been practiced should X-linkage be introduced as a modifier. Apart from the strategic approach, students should also be supported in setting up coherent arguments consisting of claims that are justified based on data. One way to do this would be to use scaffolding ( McNeill et al. , 2006 ).
Tsui and Treagust (2010) highlight that the use of human genetic pedigrees representing Mendelian problems can promote a deterministic notion of genes. To address this issue, both clear and simple pedigrees and ambiguous pedigrees should be used. It is possible to use ambiguous pedigrees (eventually showing complex traits) to generate research questions or hypotheses for further teaching; this would put a special focus on scientific inquiry. It should be possible, for example, to introduce the phenomenon of incomplete penetrance by using a pedigree that does not seem to fit with any known mode of inheritance. Stewart et al. (2005) described the use of human pedigrees in relation to blood types and achondroplasia to demonstrate that the model of simple dominance is not always adequate. These observations are the starting point for studying these “anomalous” inheritance patterns ( Stewart et al. , 2005 ). Naturally, X-linkage could be considered a modification of simple dominance in the same way ( Collins and Stewart, 1989 ). In particular, pedigrees with an unexpected gender distribution among those affected would be a good starting point here. Overall, by emphasizing reasoning and scientific inquiry, pedigree analysis can make a big contribution to the development of genetic literacy at large.
Teaching and practicing pedigree analysis should not be limited to teaching the use of content-specific concepts. Rather, the falsification strategy should also be taught and practiced. One possible way to do so would be to encourage students to use a falsification strategy with the instructions given. For example: “Try to exclude as many modes of inheritance as possible. Justify your decisions.” The use of a falsification approach could be triggered even more vigorously by further structuring and dividing the pedigree analysis into individual tasks per inheritance. For example: “Complete the following sentence. The trait is not inherited in an autosomal recessive manner because …” Moreover, it should be emphasized that a rigorous use of the falsification strategy can help check the results for plausibility and ensure that the analysis is complete. A complete analysis should provide an evaluation for all four modes of inheritance. In pedigree problems used in schools, at least two modes of inheritance can often be ruled out without doubt. Even in the case of an X-linked mode of inheritance, where no definite decision is possible, there are usually recognizable clues in favor of one mode of inheritance.
Because our research, like previous studies on pedigree analysis, did not specifically focus on teaching pedigree analysis, there are still a number of unanswered questions. In pedigree analysis, it is of primary importance to identify critical cues and to draw the right conclusions from them. In our opinion, however, it is still unclear which is the most effective way to teach students to identify these critical cues. On the one hand, there could be an ideal strategy to teach this ability; on the other hand, it could simply develop through practice. In any case, practice certainly plays an important role in pedigree analysis; therefore, students should definitely be offered a variety of pedigrees to practice with (see Timm et al. , 2020 ). Manifold practice opportunities are particularly useful if students can write down arguments for their solutions and receive individual feedback each time. This would be very time-consuming for teachers but could possibly be automated via a machine-learning approach (see Zhai et al. , 2021 ) in which students’ arguments are automatically categorized to select appropriate feedback.
Supplementary Material
Acknowledgments.
We thank Dr. Yvonne Lettmann for good advice and fruitful discussions and Marleen Gerwin for helping with the coding. This project was financially supported by the German Federal Ministry of Education and Research (BMBF) under the reference number 01PL16075. We made use of professional editing services, which was financially supported by the Interdisciplinary Center for Educational Research (IZfB) of the University of Duisburg-Essen, Germany. We acknowledge support by the Open Access Publication Fund of the University of Duisburg-Essen. Any opinions, findings, conclusions, or recommendations expressed in this publication are those of the authors and do not necessarily reflect the views of the funders. We confirm that there are no potential conflicts arising from these arrangements.
- Abraham, J. K., Perez, K. E., Price, R. M. (2014). The Dominance Concept Inventory: A tool for assessing undergraduate student alternative conceptions about dominance in Mendelian and population genetics . CBE—Life Sciences Education , 13 , 2 , 349–358. 10.1187/cbe.13-08-0160 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Aznar, M. M., Orcajo, T. I. (2005). Solving problems in genetics . International Journal of Science Education , 27 ( 1 ), 101–121. 10.1080/09500690410001673801 [ CrossRef ] [ Google Scholar ]
- Bennett, R. L. (2010). The practical guide to the genetic family history (2nd ed.). Hoboken, NJ: Wiley-Blackwell. 10.1002/9780470568248 [ CrossRef ] [ Google Scholar ]
- Bennett, R. L., Steinhaus, K. A., Uhrich, S. B., O’Sullivan, C. (1993). The need for developing standardized family pedigree nomenclature . Journal of Genetic Counseling , 2 ( 4 ), 261–273. 10.1007/BF00961575 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Bennett, R. L., Steinhaus, K. A., Uhrich, S. B., O’Sullivan, C. K., Resta, R. G., Lochner-Doyle, D., ... & Hamanishi, J. (1995). Recommendations for standardized human pedigree nomenclature . Journal of Genetic Counseling , 4 ( 4 ), 267–279. 10.1007/BF01408073 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Bennett, R. L., Steinhaus French, K., Resta, R. G., Lochner Doyle, D. (2008). Standardized human pedigree nomenclature: Update and assessment of the recommendations of the National Society of Genetic Counselors . Journal of Genetic Counseling , 17 ( 5 ), 424–433. 10.1007/s10897-008-9169-9 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Berland, L. K., McNeill, K. L. (2010). A learning progression for scientific argumentation: Understanding student work and designing supportive instructional contexts . Science Education , 94 ( 5 ), 765–793. 10.1002/sce.20402 [ CrossRef ] [ Google Scholar ]
- Cohen, J. (1960). A coefficient of agreement for nominal scales . Educational and Psychological Measurement , 20 ( 1 ), 37–46. [ Google Scholar ]
- Collins, A., Stewart, J. H. (1989). The knowledge structure of Mendelian genetics . American Biology Teacher , 51 ( 3 ), 143–149. 10.2307/4448880 [ CrossRef ] [ Google Scholar ]
- Corbett, A., Kauffman, L., Maclaren, B., Wagner, A., Jones, E. (2010). A cognitive tutor for genetics problem solving: Learning gains and student modeling . Journal of Educational Computing Research , 42 ( 2 ), 219–239. 10.2190/EC.42.2.e [ CrossRef ] [ Google Scholar ]
- DeCuir-Gunby, J. T., Marshall, P. L., McCulloch, A. W. (2011). Developing and using a codebook for the analysis of interview data: An example from a professional development research project . Field Methods , 23 ( 2 ), 136–155. 10.1177/1525822X10388468 [ CrossRef ] [ Google Scholar ]
- Freidenreich, H. B., Duncan, R. G., Shea, N. (2011). Exploring middle school students’ understanding of three conceptual models in genetics . International Journal of Science Education , 33 ( 17 ), 2323–2349. 10.1080/09500693.2010.536997 [ CrossRef ] [ Google Scholar ]
- Hackling, M. W. (1994). Application of genetics knowledge to the solution of pedigree problems . Research in Science Education , 24 ( 1 ), 147–155. 10.1007/BF02356339 [ CrossRef ] [ Google Scholar ]
- Hackling, M. W., Lawrence, J. A. (1988). Expert and novice solutions of genetic pedigree problems . Journal of Research in Science Teaching , 25 ( 7 ), 531–546. 10.1002/tea.3660250703 [ CrossRef ] [ Google Scholar ]
- Hickey, D. T., Wolfe, E. W., Kindfield, A. C. H. (2000). Assessing learning in a technology-supported genetics environment: Evidential and systemic validity issues . Educational Assessment , 6 ( 3 ), 155–196. 10.1207/S15326977EA0603_1 [ CrossRef ] [ Google Scholar ]
- Hofmann, H. (2008). Mosaic plots and their variants . In Chen, C.-H., Härdle, W., Unwin, A. (Eds.), Handbook of data visualization (pp. 617–642). Berlin, Germany: Springer. [ Google Scholar ]
- Knippels, M.-C. P. J., Waarlo, A. J., Boersma, K. T. (2005). Design criteria for learning and teaching genetics . Journal of Biological Education , 39 ( 3 ), 108–112. 10.1080/00219266.2005.9655976 [ CrossRef ] [ Google Scholar ]
- Lai, M.-L., Tsai, M.-J., Yang, F.-Y., Hsu, C.-Y., Liu, T.-C., Lee, S. W.-Y., ... & Tsai, C.-C. (2013). A review of using eye-tracking technology in exploring learning from 2000 to 2012 . Educational Research Review , 10 , 90–115. 10.1016/j.edurev.2013.10.001 [ CrossRef ] [ Google Scholar ]
- Landis, J. R., Koch, G. G. (1977). The measurement of observer agreement for categorical data . Biometrics , 33 ( 1 ), 159–174. 10.2307/2529310 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Lee, H.-S., Liu, O. L., Pallant, A., Roohr, K. C., Pryputniewicz, S., Buck, Z. E. (2014). Assessment of uncertainty-infused scientific argumentation . Journal of Research in Science Teaching , 51 ( 5 ), 581–605. 10.1002/tea.21147 [ CrossRef ] [ Google Scholar ]
- McNeill, K. L., Lizotte, D. J., Krajcik, J., Marx, R. W. (2006). Supporting students’ construction of scientific explanations by fading scaffolds in instructional materials . Journal of the Learning Sciences , 15 ( 2 ), 153–191. 10.1207/s15327809jls1502_1 [ CrossRef ] [ Google Scholar ]
- Ministry for School and Further Education of the State of North Rhine-Westphalia. (2014). Kernlehrplan für die Sekundarstufe II Gymnasium/Gesamtschule in Nordrhein-Westfalen. Biologie [Core curriculum for the upper secondary level Gymnasium/Gesamtschule in North Rhine-Westphalia. Biology] . Düsseldorf, Germany. Retrieved January 21, 2022, from https://www.schulentwicklung.nrw.de/lehrplaene/lehrplan/147/KLP_GOSt_Biologie.pdf [ Google Scholar ]
- O’Connor, C., Joffe, H. (2020). Intercoder reliability in qualitative research: Debates and practical guidelines . International Journal of Qualitative Methods , 19 , 1–13. 10.1177/1609406919899220 [ CrossRef ] [ Google Scholar ]
- Sadler, T. D., Fowler, S. R. (2006). A threshold model of content knowledge transfer for socioscientific argumentation . Science Education , 90 ( 6 ), 986–1004. 10.1002/sce.20165 [ CrossRef ] [ Google Scholar ]
- Sadler, T. D., Zeidler, D. L. (2005). The significance of content knowledge for informal reasoning regarding socioscientific issues: Applying genetics knowledge to genetic engineering issues . Science Education , 89 ( 1 ), 71–93. 10.1002/sce.20023 [ CrossRef ] [ Google Scholar ]
- Sandoval, W. A., Millwood, K. A. (2005). The quality of students’ use of evidence in written scientific explanations . Cognition and Instruction , 23 ( 1 ), 23–55. 10.1207/s1532690xci2301_2 [ CrossRef ] [ Google Scholar ]
- Schmiemann, P., Nehm, R. H., Tornabene, R. E. (2017). Assessment of genetics understanding: Under what conditions do situational features have an impact on measures? Science & Education , 26 ( 10 ), 1161–1191. 10.1007/s11191-017-9925-z [ CrossRef ] [ Google Scholar ]
- Shea, N. A., Duncan, R. G., Stephenson, C. (2015). A tri-part model for genetics literacy: Exploring undergraduate student reasoning about authentic genetics dilemmas . Research in Science Education , 45 ( 4 ), 485–507. 10.1007/s11165-014-9433-y [ CrossRef ] [ Google Scholar ]
- Smith, M. U. (1988). Successful and unsuccessful problem solving in classical genetic pedigrees . Journal of Research in Science Teaching , 25 ( 6 ), 411–433. 10.1002/tea.3660250602 [ CrossRef ] [ Google Scholar ]
- Smith, M. U., Good, R. (1984). Problem solving and classical genetics: Successful versus unsuccessful performance . Journal of Research in Science Teaching , 21 ( 9 ), 895–912. 10.1002/tea.3660210905 [ CrossRef ] [ Google Scholar ]
- Songer, N. B., Kelcey, B., Gotwals, A. W. (2009). How and when does complex reasoning occur? Empirically driven development of a learning progression focused on complex reasoning about biodiversity . Journal of Research in Science Teaching , 46 ( 6 ), 610–631. 10.1002/tea.20313 [ CrossRef ] [ Google Scholar ]
- Standing Conference of the Ministers of Education and Cultural Affairs of the Länder in the Federal Republic of Germany. (2005). Bildungsstandards im Fach Biologie für den Mittleren Schulabschluss [Educational standards in biology for the intermediate school leaving certificate] . München, Germany: Luchterhand. Retrieved January 21, 2022, from https://www.kmk.org/fileadmin/Dateien/veroeffentlichungen_beschluesse/2004/2004_12_16-Bildungsstandards-Biologie.pdf [ Google Scholar ]
- Stewart, J. (1988). Potential learning outcomes from solving genetics problems: A typology of problems . Science Education , 72 ( 2 ), 237–254. 10.1002/sce.3730720211 [ CrossRef ] [ Google Scholar ]
- Stewart, J., Cartier, J. L., Passmore, C. M. (2005). Developing understanding through model-based inquiry . In National Research Council, How students learn: History, mathematics, and science in the classroom (pp. 515–565). Washington, DC: National Academies Press. [ Google Scholar ]
- Surmann, J. (2017). Teilautomatisierte Entwicklung und Analyse humangenetischer Stammbäume [Semi-automated development and analysis of human genetic pedigrees] (Unpublished master’s thesis). Germany: University of Duisburg-Essen. [ Google Scholar ]
- Therneau, T. M., Sinnwell, J. (2015). Kinship2: Pedigree functions (R package version 1.6.4) . Retrieved March 7, 2018, from https://CRAN.R-project.org/package=kinship2
- Timm, J., Otto, B., Schramm, T., Striewe, M., Schmiemann, P., Goedicke, M. (2020). Technical aspects of automated item generation for blended learning environments in biology . I-Com , 19 ( 1 ), 3–15. 10.1515/icom-2020-0001 [ CrossRef ] [ Google Scholar ]
- Toulmin, S. E. (2003). The uses of argument (Updated ed.). Cambridge, England: Cambridge University Press. Retrieved from www.netLibrary.com/urlapi.asp?action=summary&v=1&bookid=120780 [ Google Scholar ]
- Tsui, C., Treagust, D. (2010). Evaluating secondary students’ scientific reasoning in genetics using a two-tier diagnostic instrument . International Journal of Science Education , 32 ( 8 ), 1073–1098. 10.1080/09500690902951429 [ CrossRef ] [ Google Scholar ]
- VERBI Software. (2018). MAXQDA 2018: The art of data analysis. Berlin .
- von Aufschnaiter, C., Erduran, S., Osborne, J., Simon, S. (2008). Arguing to learn and learning to argue: Case studies of how students’ argumentation relates to their scientific knowledge . Journal of Research in Science Teaching , 45 ( 1 ), 101–131. 10.1002/tea.20213 [ CrossRef ] [ Google Scholar ]
- Wools, K. (2018). Strategien von Lernenden bei der Analyse von Familienstammbäumen [Strategies of learners in family tree analysis] (Unpublished master’s thesis). Germany: University of Duisburg-Essen. [ Google Scholar ]
- Zhai, X., Shi, L., Nehm, R. H. (2021). A meta-analysis of machine learning-based science assessments: Factors impacting machine-human score agreements . Journal of Science Education and Technology , 30 ( 3 ), 361–379. 10.1007/s10956-020-09875-z [ CrossRef ] [ Google Scholar ]
- Undergraduate
- High School
- Architecture
- American History
- Asian History
- Antique Literature
- American Literature
- Asian Literature
- Classic English Literature
- World Literature
- Creative Writing
- Linguistics
- Criminal Justice
- Legal Issues
- Anthropology
- Archaeology
- Political Science
- World Affairs
- African-American Studies
- East European Studies
- Latin-American Studies
- Native-American Studies
- West European Studies
- Family and Consumer Science
- Social Issues
- Women and Gender Studies
- Social Work
- Natural Sciences
- Pharmacology
- Earth science
- Agriculture
- Agricultural Studies
- Computer Science
- IT Management
- Mathematics
- Investments
- Engineering and Technology
- Engineering
- Aeronautics
- Medicine and Health
- Alternative Medicine
- Communications and Media
- Advertising
- Communication Strategies
- Public Relations
- Educational Theories
- Teacher's Career
- Chicago/Turabian
- Company Analysis
- Education Theories
- Shakespeare
- Canadian Studies
- Food Safety
- Relation of Global Warming and Extreme Weather Condition
- Movie Review
- Admission Essay
- Annotated Bibliography
- Application Essay
- Article Critique
- Article Review
- Article Writing
- Book Review
- Business Plan
- Business Proposal
- Capstone Project
- Cover Letter
- Creative Essay
- Dissertation
- Dissertation - Abstract
- Dissertation - Conclusion
- Dissertation - Discussion
- Dissertation - Hypothesis
- Dissertation - Introduction
Dissertation - Literature
- Dissertation - Methodology
- Dissertation - Results
- GCSE Coursework
- Grant Proposal
- Marketing Plan
- Multiple Choice Quiz
- Personal Statement
- Power Point Presentation
- Power Point Presentation With Speaker Notes
- Questionnaire
- Reaction Paper
Research Paper
- Research Proposal
- SWOT analysis
- Thesis Paper
- Online Quiz
- Literature Review
- Movie Analysis
- Statistics problem
- Math Problem
- All papers examples
- How It Works
- Money Back Policy
- Terms of Use
- Privacy Policy
- We Are Hiring
The Family Pedigree Analysis, Research Paper Example
Pages: 2
Words: 567
Hire a Writer for Custom Research Paper
Use 10% Off Discount: "custom10" in 1 Click 👇
You are free to use it as an inspiration or a source for your own work.
After carefully analyzing my family pedigree, there are two important trends to notice with regards to diseases passed along throughout the family. The first disease of note is on my parental side by which there are chronic cases of Stroke or Brain Attack. In the case of my uncle, it is likely that this was caused by a developed brain tumor. However, a more interesting disease on my maternal side comes from the chronic issue of high cholesterol. As mentioned in a previous lab assignment, current research shows that high cholesterol can not only be inherited, but it can also lead to inherited disorders such as familial Hyperlipidemia and Hypercholesterolemia.
Due to the extent of the low and uncommon prevalence of high cholesterol in my family, it appears as though the occurrence of high cholesterol follows an autosomal recessive mode of inheritance. It currently has only proven to be present in my maternal family starting with my grandfather and most recently prevalence has appeared in my mother. According to BiologyReference.com (2008), “parents of affected individuals are unaffected carriers” whenever an autosomal recessive mode of inheritance exists for a disease. Furthermore, “each of the parent’s offspring has a 25 percent chance of inheriting the illness and a 75 percent chance of being unaffected” (BiologyReference.com, 2008). My mother was one of six children from my grandparents as was the only child to share high cholesterol with my grandfather. It would appears that the autosomal recessive inheritance is correct in this case because of this fact.
In this scenario, it does not appear that the mutation is a compound heterozygous mutation because there is no prevalence or previous history of high cholesterol in my paternal family. Therefore, the mutation must be heterozygous and most of the infected individuals are unaffected carriers of the gene and there are no visible signs of high cholesterol. With this in mind, this could point to the fact that not only am I a potential carrier, but so too may be my siblings. If I am an unaffected carrier, it is unclear whether the mutation will be passed on to my children and whether they will be homozygous or heterozygous for high cholesterol.
Perhaps a stronger knowledge of my pedigree would be useful to understand whether high cholesterol in my maternal family was originally autosomal recessive or dominant. Because only my grandfather had high cholesterol, not my grandmother, evidence appears to show that the inheritance was autosomal recessive, although there is the possibility that this could have mutated differently in my family. The main problem associated with compiling my pedigree was the lack of attention that my family has paid to such diseases because they are often rare in the family. Many of my siblings and parents’ siblings could not recall their parents’ health histories. My pedigree accuracy was slightly compromised because of this. There may be additional health issues within the family that we are not aware of. I believe that I can tell my children that their chances of high cholesterol are fairly low. They may turn out to be unaffected carriers and show no signs of high cholesterol, but they should still maintain a healthy and active lifestyle to counter the issues associated with high cholesterol just to be safe.
BiologyReference.com. (2008). Pedigrees and modes of inheritance . Retrieved from http://www.biologyreference.com/Oc-Ph/Pedigrees-and-Modes-of-Inheritance.html
Rodriguez, D. (2009, June 17). High cholesterol: are your genes to blame?. Everyday Health , Retrieved from http://www.everydayhealth.com/high-cholesterol/high-cholesterol-are-genes-to-blame.aspx
Stuck with your Research Paper?
Get in touch with one of our experts for instant help!
Rationale Objectives, Dissertation - Literature Example
Human Resources Management as a Discipline, Essay Example
Time is precious
don’t waste it!
Plagiarism-free guarantee
Privacy guarantee
Secure checkout
Money back guarantee

Related Research Paper Samples & Examples
The risk of teenagers smoking, research paper example.
Pages: 11
Words: 3102
Impacts on Patients and Healthcare Workers in Canada, Research Paper Example
Pages: 7
Words: 1839
Death by Neurological Criteria, Research Paper Example
Words: 2028
Ethical Considerations in End-Of-Life Care, Research Paper Example
Pages: 5
Words: 1391
Ethical Dilemmas in Brain Death, Research Paper Example
Words: 2005
Politics of Difference and the Case of School Uniforms, Research Paper Example
Words: 631

- Previous Article
- Next Article
Introduction
Pedigree construction, modes of inheritance, interpreting pedigrees, charles b. davenport, problems with davenport’s pedigrees analysis, eugenics old & new, the misuse of pedigree analysis in the eugenics movement.
MARK SHOTWELL is an Associate Professor of Biology at Slippery Rock University, Slippery Rock, PA 16057; e-mail: [email protected] .
- Split-Screen
- Article contents
- Figures & tables
- Supplementary Data
- Peer Review
- Open the PDF for in another window
- Guest Access
- Get Permissions
- Cite Icon Cite
- Search Site
Mark Shotwell; The Misuse of Pedigree Analysis in the Eugenics Movement. The American Biology Teacher 1 February 2021; 83 (2): 80–88. doi: https://doi.org/10.1525/abt.2021.83.2.80
Download citation file:
- Ris (Zotero)
- Reference Manager
Pedigree analysis has long been an essential tool in human genetics as well as a staple of genetics education. Students of genetics might be surprised to learn that human pedigrees were first popularized in the United States by proponents of eugenics, the pseudoscientific social movement aimed at improving the genetic quality of the human race. Notably, the influential eugenicist Charles B. Davenport employed pedigree charts to support his belief that not only were such medical conditions as Huntington disease and albinism inherited in a simple Mendelian fashion, but so too were such characteristics as alcoholism, criminality, and “feeblemindedness.” We now see the flaws in Davenport’s pedigree analysis, but at the time, it was the latest scientific advance. The misuse of pedigree analysis during the eugenics era may serve as a cautionary tale for those who are now harnessing the latest genetic technologies to solve complex problems.
Humans make terrible subjects for genetic studies. That’s because the primary experimental tool of genetics, the controlled cross, is simply not possible for humans. There is no such thing as a highly homozygous “true-breeding line” of humans, and even if there were, it would be impossible (or at the very least highly unethical) to compel two people to mate with each other to reveal the pattern of inheritance of a particular trait. Almost since the dawn of genetics at the turn of the 20th century, therefore, human geneticists have relied on a much less powerful (but at least feasible) method, the analysis of pedigree charts.
“Eugenicists employed pedigrees to bolster their claims that not only were physical traits strictly inherited as simple Mendelian characters, but so too were mental, emotional, and behavioral characteristics.”
Pedigree charts are branching diagrams that depict the appearance of a trait in a family ( Resta, 1993 ). If the family is large enough, comprising several generations with multiple affected persons, it may be possible to infer how the trait is inherited, whether the responsible gene resides on the X chromosome or an autosome, and whether it is dominant or recessive.
Genealogical charts were drawn for centuries before they became so important in human genetics. At first, they were simply graphical representations of family relationships. The name is derived from the Middle French pié de grue , literally meaning “foot of crane,” because some early diagrams contained so many branching diagonal lines that they resembled a bird’s foot. One of the earliest surviving pedigrees showed the inheritance of hemophilia A, an X-linked recessive condition, in the royal families of Europe.
Pedigrees have grown in sophistication in recent decades, and may now incorporate information from genetic testing as specific as single-nucleotide differences in genes. But their basic construction has changed little since the first decades of the 20th century. What most students of genetics are unaware of, however, is that pedigree analysis was developed in this country by proponents of eugenics, the pseudoscientific social movement with the aim of controlling human heredity ( Resta, 1993 ). Eugenicists employed pedigrees to bolster their claims that not only were physical traits strictly inherited as simple Mendelian characters, but so too were mental, emotional, and behavioral characteristics. As we will see, their analysis was deeply flawed, based on slipshod data collection, unsupported assumptions, and circular reasoning.
Pedigree construction became standardized in the United States in 1911 with the publication of the book Heredity in Relation to Eugenics by the leading American geneticist and eugenicist Charles B. Davenport ( Davenport, 1911 ). Until that time, pedigree charts used a variety of symbols; from that time onward, almost all were made with the symbols shown in Figure 1 (at least in the United States).

Standard symbols used in pedigree charts.
Davenport’s pedigree of a family with Huntington disease illustrates the key features of pedigree construction ( Figure 2 ). Females are indicated with circles and males with squares. (Previously, ♀ and ♂ were used for female and male, respectively.) Affected persons are denoted by shaded symbols. The generations each occupy a separate line, with the most ancestral at the top, and are numbered with Roman numerals. Individuals within a generation are numbered left to right with Arabic numerals. Each person in the pedigree may thus be identified by a Roman numeral and an Arabic numeral (e.g., II-3). A single line connects a female and male to represent a mating, and a vertical line of descent drops from the mating line to the sibship line, which connects the offspring in their order of birth.

Davenport’s pedigree of a family with Huntington disease ( Davenport, 1911 , p. 103).
The goal when examining a pedigree is to infer the mode of inheritance of the trait. Does the pattern of appearance of the trait suggest autosomal dominant, autosomal recessive, X-linked dominant, or X-linked recessive inheritance? The first step is to determine whether there are any skipped generations; that is, are there any instances of two unaffected parents with an affected child? For traits determined by a recessive allele, it is possible, even common, for two unaffected parents to have an affected child. Both parents must be heterozygous, however ( Figure 3 ). If the trait is determined by a dominant allele, two unaffected parents cannot have an affected child. One or the other of the parents would have to carry the dominant allele for a child to be affected ( Figure 4 ). Skipped generations are thus diagnostic of a recessive mode of inheritance. The absence of skipped generations, when every affected person has at least one affected parent, is evidence for dominant inheritance.

A skipped generation is consistent with autosomal recessive inheritance.

A skipped generation is inconsistent with autosomal dominant inheritance.
Once dominant vs. recessive inheritance has been established, the next task is to decide whether the responsible gene resides on an autosome or on the X chromosome. We’ll start with the recessive mode. For the X-linked recessive mode of inheritance, females with genotypes AA and Aa are unaffected, and only genotype aa is affected. In males, who have only one X chromosome and therefore are hemizygous for all X-linked genes, A Y is unaffected and a Y is affected. The genotypes and the associated pedigree symbols are shown in Figure 5 .

Pedigree symbols used for X-linked recessive inheritance.
Consider a mating between a heterozygous unaffected female (or “carrier female”) and a hemizygous dominant unaffected male ( Figure 6 ). The possible outcomes of this mating are given in the Punnett square in Figure 7 . Note that this is an example of two unaffected parents having an affected child, a “skipped generation,” the diagnostic element of recessive inheritance. Note further that the expected phenotype ratio is three unaffected to one affected, but all of the affected are male. In other words, the recessive phenotype co-segregates with sex.

Mating between a “carrier female” and an unaffected male.

Expected outcomes of the mating in Figure 7.
The appearance of the phenotype usually (but not always) skips generations. That is, affected persons almost always have parents who are both unaffected.
The phenotype appears predominantly in males. For a rare condition, affected females will almost never be found.
The pedigree depicting the occurrence of hemophilia in the royal families of Europe (mentioned above; the pedigree can easily be found online) shows these two characteristic features. So does the pedigree in Figure 8 , which depicts the appearance of Duchenne muscular dystrophy in a small three-generation family ( Bundey, 1978 ).

Pedigree showing the appearance of Duchenne muscular dystrophy, an X-linked recessive condition.
If there are skipped generations in a pedigree, but the trait is not far more common in males, it may be concluded that the mode is autosomal recessive. Davenport’s pedigree of a family with astigmatism shows the characteristic features of autosomal recessive inheritance ( Figure 9 ).

Davenport’s pedigree of a family with astigmatism ( Davenport, 1911 , p. 123).
Of the four possible modes of inheritance, X-linked dominant is by far the least common. Only a very few X-linked dominant conditions are known; incontinentia pigmenti and Rett syndrome are two on a very short list. Given its rarity, X-linked dominant inheritance will not be discussed here.
If there are no skipped generations (i.e., no instances of two unaffected parents with at least one affected child), autosomal dominant inheritance may be assumed. (X-linked dominant inheritance can be ruled out by the appearance of either an affected father with an affected son or an affected father with an unaffected daughter.)
If the phenotype is much more frequent in males than in females (or appears only in males), X-linked recessive inheritance may be assumed. All unaffected females must have at least one dominant allele ( A –). The affected males are hemizygous recessive ( a Y) and the unaffected males are hemizygous dominant ( A Y).
If the trait is not much more frequent in males than in females, autosomal recessive inheritance is assumed. Everyone unaffected must have at least one dominant allele ( A –), and all the affected persons are homozygous recessive ( aa ).
Consider the pedigree in Figure 10 , showing the appearance of alkaptonuria, a defect in the gene encoding the enzyme homogentisic acid oxygenase ( Cuthbert, 1923 ). The first question we ask ourselves about this pedigree is whether there are any skipped generations. The answer to this question is yes: III-1 and III-2 are both unaffected and have three affected children (IV-1, IV-3, and IV4). We therefore conclude that alkaptonuria is a “recessive trait.”

Pedigree showing the appearance of alkaptonuria, an autosomal recessive condition.
Our second question is whether the phenotype is much more common in males than in females. In this case, there are two affected males (IV-1 and IV-3) and one affected female (IV-4). A ratio of two affected males to one affected female is far too low for this to be X-linked recessive. We thus conclude that the mode of inheritance is autosomal recessive.
Alkaptonuria (black urine disease) was the first so-called inborn error of metabolism described by Archibald Garrod ( Garrod, 1902 ). Garrod himself suggested that alkaptonuria was a recessive condition (based on his discussions with William Bateson). Garrod also noted that 60% of the cases of alkaptonuria he had identified appeared in the children of first-cousin marriages. This is another hallmark of the autosomal recessive mode of inheritance, a higher incidence in the offspring of consanguineous matings, most commonly between first and second cousins. Such matings are indicated by a double mating line, as shown in Figure 10 .
Charles Benedict Davenport ( Figure 11 ) was a Harvard-educated zoologist who helped extend Mendelian principles to animals, including poultry, mice, and horses, in the first decade of the 20th century. During that time, he also carried out a series of studies of eye color, hair form, and hair color in humans, and proposed that skin color is determined by two interacting genes. Consequently, Davenport may rightly be considered the first true human geneticist.

Charles B. Davenport, 1866–1944. Photo credit: http://campus.udayton.edu/~hume/Eugenics/eugenics.htm (accessed February 7, 2014). Used by permission of the American Philosophical Society.
With equal justification, Davenport may also be regarded as the leading eugenicist in the United States. Eugenics was a pseudoscientific social movement whose goal was to improve the hereditary quality of the human race by controlling breeding, encouraging the hereditarily “superior” to have more children and discouraging (or preventing altogether) the reproduction of the genetically “inferior.” Eugenics originated in the writings of the English polymath Francis Galton (Charles Darwin’s half-cousin) and flourished in the first three decades of the 20th century, stimulated by the rediscovery of Mendel’s principles of heredity in 1900. Eugenics programs sprung up throughout Europe, Scandinavia, and South America, in addition to England and the United States. In the United States, eugenics found widespread favor, supported by prominent authors (e.g., H. G. Wells), journalists (e.g., Albert E. Wiggam), industrialists (e.g., John Harvey Kellogg), inventors (e.g., Alexander Graham Bell), birth control advocates (e.g., Margaret Sanger), psychologists (e.g., Henry H. Goddard), religious leaders (e.g., William R. Inge), and politicians (e.g., Theodore Roosevelt). It was also promoted by the leading geneticists of the day, including R. A. Fisher in England, Erwin Baur in Germany, Herman Nilsson-Ehle in Sweden, and Edward M. East in the United States, among many others ( Paul & Spencer, 1995 ). For a time, eugenics was backed by the pioneering geneticist Thomas Hunt Morgan, whose work with fruit flies first showed that genes reside on chromosomes. Though lukewarm in his support, Morgan did serve as a founding scientific director of the Eugenics Record Office ( Lombardo, 2001 ). He soon grew disillusioned with the methods of the eugenics movement, criticizing them as reckless and unreliable to Davenport ( Ludmerer, 1972 , pp. 82–83). In 1915 he resigned in protest from the American Breeders’ Association ( Kevles, 1985 , p. 122), which, under Davenport’s influence, had begun strongly promoting eugenics ( Kimmelman, 1983 , p. 185).
Davenport published influential articles and books, edited journals, founded professional societies, and organized meetings of like-minded enthusiasts of scientific breeding in humans. His most significant contribution to the field, however, was acquiring generous funding to promote eugenics. In 1910 he persuaded Mary Harriman, the widow of the railroad baron E. H. Harriman, to contribute a large sum of money (equivalent to half a million of today’s dollars annually) to establish the Eugenics Record Office at Cold Spring Harbor, New York ( Figure 12 ). Later financial support came from oil magnate John D. Rockefeller Jr. and the Carnegie Institution of Washington. The ERO had a twofold purpose: (1) to advance research in human genetics and, in particular, to elucidate the manner of inheritance of specific human traits; and (2) to educate the public about eugenics by disseminating findings and supporting nationwide eugenic education efforts ( Allen, 1986 ). More than 250 fieldworkers were trained at the ERO (the majority of them young college-educated women) to administer mental tests, record physical measurements, and recognize insanity, criminality, epilepsy, and other conditions. Their training included doing genetic crosses in maize so that they would see Mendelian patterns of inheritance for themselves, patterns they were expected to find in the families they studied ( Wilson, 2002 ). They fanned out to state hospitals, insane asylums, poorhouses, and reformatories, as well as people’s homes, ultimately amassing hundreds of thousands of pedigree charts, family histories, and other hereditary data, which were catalogued and stored at the ERO. The ERO’s pedigrees collected by Davenport and those constructed by the psychologist Henry H. Goddard were disseminated widely, appearing in hundreds of biology textbooks ( Largent, 2008 , p. 129) as well as in publications intended for a more general audience. Ostensibly serving an educational purpose, these pedigree charts became tools of propaganda, persuading the public of the dangers of hereditary degeneration and the urgent need for legislative remedies ( Lombardo, 2001 ).

The Eugenics Record Office archives room. Photo credit: www.eugenicsarchive.org .
As a staunch Mendelian, Davenport considered every human characteristic to be controlled by a single gene with dominant and recessive alleles whose expression is not fundamentally altered by the environment (to be contrasted with a multifactorial trait, whose expression is affected by alleles of several – or even many – genes as well as by environmental conditions). Hair color and texture, eye color, temperament, mathematical ability, alcoholism, musical talent, muscular strength, nervousness, and “feeblemindedness” – all were equally simple Mendelian characters to Davenport, each determined by a single gene with a dominant and a recessive allele.
The first fruits of the ERO’s data-collection effort appeared in Heredity in Relation to Eugenics ( Davenport, 1911 ). It contained just about all there was to know about human genetics at the time and included more than 150 pedigree charts, for everything from cataracts to criminality, from epilepsy to eroticism, from ichthyosis to insanity. Some of Davenport’s pedigrees did, in fact, reveal the mode of inheritance of a trait. He correctly deduced that not only was Huntington’s disease inherited as a dominant trait but so were achondroplasia and polydactyly. Albinism was found to be recessively inherited. His pedigree for albinism is reproduced in Figure 13 . Examination of this pedigree reveals multiple skipped generations, a substantial proportion of affected females (9 out of 19), and several consanguineous matings, all characteristic of the autosomal recessive mode of inheritance. Note that this pedigree shows females and males not as circles and squares, but as the symbols ♀ and ♂, respectively, a convention followed by English eugenicists such as Karl Pearson ( Resta, 1993 ).

Pedigree chart of an albino family (♀ = female, ♂ = male, ○ = sex unknown). Filled circles represent albinos. The letters B, P, R, and W represent the four common surnames in this highly inbred community.
Many of the pedigree charts Davenport presented in Heredity in Relation to Eugenics were of little value, however. Figure 14 , showing the appearance of criminality in a family, serves to illustrate some of the problems in Davenport’s analysis. From the skipped generation in this pedigree, Davenport concluded that criminality is inherited as a recessive trait, “like most neuroses.” Upon closer examination, we find the basis for this inference very shaky indeed. What was the evidence for criminality in this family? Davenport related that the first affected person (I-3) was “a western desperado, drank hard and was involved in a murder.” His grandson (person II-2) was reported to have hitched a ride on a train at age three, run away from reform school 13 times, lied habitually, reneged on debts, and been convicted of burglary.

Davenport’s pedigree showing the appearance of criminality.
In Heredity in Relation to Eugenics , Davenport wrote that many other traits were mainly, if not entirely, inherited in a recessive mode, including musical, artistic, and literary abilities (p. 61), “bodily energy” (p. 93), epilepsy (p. 72), insanity (“neuropathic taint,” p. 77), alcoholism (“a strong hereditary bias toward alcohol,” p. 83), feeblemindedness (pp. 65–72), and sexual immorality (pp. 90–92).
In a later book, Davenport presented exhaustive evidence that the trait thalassophilia (literally “love of the sea”), a type of maritime wanderlust, was inherited in families of naval officers ( Davenport & Scudder, 1919 ). One such family was that of British Vice-Admiral Cuthbert Collingwood, the hero of the Battle of Trafalgar in 1805 ( Figure 15 ). On the basis of this and many additional pedigrees, Davenport concluded that thalassophilia was recessive and sex-limited because it often skipped a generation and occurred only in males. (His research had failed to uncover any naval officers who were women.)

Davenport’s pedigree showing the appearance of “thalassophilia.”
There are many problems with the pedigrees Charles Davenport constructed and interpreted using data collected by ERO fieldworkers. First, although many of the traits he considered were straightforward conditions such as albinism, polydactyly, and color blindness, many were complex and ill-defined, like alcoholism, musical ability, pauperism, mechanical skill, general mental ability, nervousness, and, of course, “feeblemindedness.” For pedigree analysis to be successful, the trait under study must be well defined and easily discriminated.
Second, the fact that a trait runs in a family does not prove that the trait has a genetic basis. It is not uncommon for traits that at first appear to be inherited to be found to have an environmental basis. (Such an environmentally caused phenotype that mimics a genetic condition is called a phenocopy.) Multiple occurrences of a disease in a family may be due to a bacterial or viral infection, a nutritional deficiency, or the exposure to an environmental toxin. It is important to remember that just because a trait is familial (i.e., running in a family), that does not necessarily mean it is genetic.
The trait must occur more frequently among the genetic relatives of an affected person than in the general population.
The trait must not spread to unrelated persons exposed to similar environmental conditions.
Another indicator is when identical twins (who have 100% of their genes in common) share the phenotype more often than nonidentical (fraternal) twins (who share 50% of their genes).
A third mistake Davenport and others made is assuming that even those traits with a substantial genetic foundation were simple Mendelian characters. In other words, he believed that each trait was controlled by a single gene with a dominant allele and a recessive allele, and that the genetic cause was the same in every family. For example, Davenport presupposed that polydactyly, which he showed to be a dominant condition, had the same underlying genetic basis in every family in which it occurred. In fact, it is now known to be caused by mutations in at least six different genes ( PAPA1 , PAPA2 , PAPA6 , SRPS2A , PPD2 , and SRPS3 ) (Online Mendelian Inheritance in Man), most of which are dominant but incompletely penetrant. This is an example of genetic heterogeneity, when the same phenotype may result from different genotypes. It is well known that mutations in the BRCA1 and BRCA2 genes increase a woman’s risk of developing breast cancer, but at least 11 other genes also raise breast cancer susceptibility when mutated ( McClellan & King, 2010 ).
An even more extreme case is inherited deafness. At least 120 genes have been identified that, when mutated, result in deafness ( Nance, 2003 ). The inheritance pattern may be autosomal dominant (i.e., gene GJB6 ), autosomal recessive (i.e., gene GJB2 ), or X-linked recessive (i.e., gene POU3F4 ). Hearing loss may even result from mutations in genes residing not on a chromosome but in the DNA of the mitochondrion ( Kokotas et al., 2007 ).
There were also many problems in the way the ERO fieldworkers collected the data Davenport analyzed. Often the information gathered was subjective, based on cursory observation and guesswork by the poorly trained fieldworkers. Even worse, fieldworkers commonly made records of people they had never met, because they were either geographically distant or no longer living. These records were based on unreliable personal recollections of family members and published sources of questionable veracity.
Compounding these issues was that many of Davenport’s pedigrees were quite small, comprising a dozen or so persons in three, or sometimes only two, generations, with many family members unaccounted for. Figure 16 is one example – a pedigree showing “heart trouble.” Based on this one pedigree, the mode of inheritance of “heart trouble” could be autosomal dominant, autosomal recessive, or X-linked dominant. Only X-linked recessive is ruled out. Such small families make it difficult, if not impossible, to discern the mode of inheritance of a trait.

Davenport’s pedigree showing the appearance of “heart trouble.”
Yet another problem was that Davenport used anecdotes to support his hereditarian claims. To him, the fact that the three Brontë sisters (Charlotte, Emily, and Anne) had each written a famous book was strong evidence of the hereditary nature of literary ability. He likewise took the similar professional accomplishments of John Roebling, the designer of the Brooklyn Bridge, and his sons, who finished the bridge and later supplied cable for the Golden Gate Bridge, as evidence that mechanical skill is inherited. Anecdotes, no matter how suggestive, do not count as scientific evidence.
Finally, Davenport and other eugenicists used circular reasoning in their analysis of human pedigrees. Here’s how it worked: First, a pedigree was produced that purported to show that a particular trait was inherited in a simple Mendelian fashion with a clear-cut mode of inheritance. Next, when this trait was found in another family, it was assumed that it displayed the same straightforward mechanism of inheritance as in that first pedigree. This new information was interpreted in such a way as to fit in this simple model, even when it plainly did not. This data manipulation lent credence to the original pedigree analysis, and elevated its conclusions to “scientific fact.”
Eventually, the flaws in the pedigree data gathered by ERO fieldworkers and analyzed by Charles Davenport became apparent. After reviewing the ERO in 1935, the year after Davenport’s retirement, a scientific committee concluded that this information was of little scientific value ( Kevles, 1985 , p. 199). The ERO was quietly shut down four years later, its influence having all but evaporated ( Paul, 1998 , p. 120).
The eugenics movement did not end when the Eugenics Record Office closed, however. As is well known, the most extreme expression of eugenics took place in Nazi Germany. Eugenics started slowly in German and for many years lagged behind the movements in England and the United States. German eugenicists monitored the developments in these countries closely and maintained close ties with such prominent American figures as Charles Davenport; his deputy, Harry Laughlin; the anthropologist Clarence G. Campbell; and, especially, the agriculturist and eugenicist Paul Popenoe, from whom they gained inspiration ( Kühl, 1994 , p. 19).
One who followed American eugenics closely was future chancellor Adolf Hitler. A mere two months after he came to power in 1933, the Law for the Prevention of Genetically Diseased Progeny was enacted. Patterned on statutes in California and other states, which were themselves based on Laughlin’s Model Eugenical Sterilization Law of 1922 , it mandated the sterilization of those with “feeblemindedness,” mental illness, epilepsy, hereditary blindness and deafness, Huntington’s disease, alcoholism, and physical deformities ( Kühl, 1994 , p. 39). In the first three years the law was in effect, 225,000 people were sterilized ( Kevles, 1985 , p. 117). Later, its reach was extended to the “socially feebleminded,” as determined by Hereditary Health Courts.
As the Third Reich gained momentum, German eugenics took ever more sinister turns, as it became subsumed by Nazi racial policies. Forced sterilizations all but ceased after 1939, replaced by a program of euthanasia. The first to die in the gas chambers were mental patients, followed by homosexuals, other social and political outcasts, about 500,000 Gypsies, and, most horrifically, 6 million Jews.
The first published histories of the eugenics movement suggested that advances in genetics gradually eroded the scientific foundation of the eugenic program, exposing the movement’s leaders to ridicule. This was accompanied by a waning of interest in eugenics during the Great Depression of the 1930s when the country had more pressing concerns than controlling human heredity. The movement was finally laid to rest when the atrocities of the Nazis came to light at the end of World War II. More recent scholarship has challenged this narrative ( Paul, 2016 ).
Compulsory sterilization of the “unfit,” the blunt instrument of negative eugenics, did not stop when the original eugenics movement ran out of steam and the ERO shut down. In fact, eugenic sterilizations increased during the depression of the 1930s as new state laws were enacted and older ones were more vigorously enforced. Sterilizations peaked in 1933 and did not begin declining until 1944 ( Largent, 2008 , p. 77). Coerced sterilizations continued to be performed for more than two decades after the end of World War II, although they were increasingly targeted at the poor and ethnic minorities rather than the “feebleminded” ( Largent, 2008 ).
At the same time, many so-called reform eugenicists began making common cause with birth-control advocates in the United States, England, and elsewhere. Their goal in providing information and distributing contraceptive devices was not so much granting women control over their reproductive lives as it was reducing the fertility of the poor ( Klausen & Bashford, 2012 ).
In the years just before and after World War II, eugenically minded geneticists helped develop a new medical specialty, which became known as genetic counseling. As with Davenport thirty years earlier, the pioneers in the field relied heavily on pedigree construction and analysis, although they were careful to confine themselves to conditions with a clear hereditary basis ( Largent, 2008 ). A hallmark of genetic counseling today is the principle of nondirectiveness; counselors present information and outline options to couples but do not recommend a course of action. But in the formative days of the profession, counselors took into account not only the welfare of the couples seeking assistance but also the future hereditary health of the entire population ( Resta, 1997 ). These early hereditary clinics were thus intended to have a role in shaping human evolution by preventing the occurrence of inherited disease ( Paul, 1998 , pp. 125–127; Comfort, 2012 , p. 119).
Interestingly, Davenport’s Heredity in Relation to Eugenics may be considered the first handbook on genetic counseling ( Reilly, 2008 , p. 160). In it, he advised those with a family history of an inherited disorder not to have children (a practice that today we would call directive counseling). He recommended against marriages between first cousins and other close relatives and went so far as to suggest that sisters of hemophiliacs should opt to remain childless (see Davenport, 1911 , pp. 118 and 157).
It is now clear that eugenics did not dwindle in the 1930s and disappear altogether at the end of World War II. Rather, it was reconceived, adapting to new developments in biology and medicine. Along the way, it was rebranded, shedding the unpalatable label “eugenics” in favor of such terms as “medical genetics” and “social biology,” all the while adhering to many of the goals of the original movement ( Paul, 2016 ).
More than a century after Charles Davenport standardized the construction of pedigree charts, pedigrees remain an essential tool in human genetics. But in recent decades, a host of genetic and reproductive technologies have been invented and perfected, including in vitro fertilization, preimplantation genetic testing, gene therapies, stem cell treatments, and, most recently, CRISPR gene editing. These technologies make possible a new eugenics of a scope and power unimaginable to Davenport. It will be driven not by propaganda campaigns and state-directed programs, but by consumer preferences stoked by a vast biotechnology industry. It will be, in other words, not an authoritarian eugenics but an individualistic eugenics.
Few would dispute that modern genetic technologies hold enormous promise for the diagnosis, treatment, and prevention of inherited disease and that this promise has only partially been realized. But the use of these technologies raises many thorny questions (see, e.g., Baylis, 2019 ). Can we harness the powerful new genetic tools to cure disease without engendering a new era of eugenics ( Comfort, 2018 )? Or is a 21st-century eugenics inevitable? If it is, will its benefits be equitably distributed throughout society, or will only the wealthy stand to gain, along with the industry that caters to their needs? Will genetic knowledge be used more wisely than it was a century ago, when pedigree charts served as the “scientific” justification for discrimination against the disadvantaged, extending so far as institutionalization and sexual sterilization? Have we learned the lessons of the original eugenics movement well enough to navigate the ethical waters that lie ahead? Or will future generations judge us for our ethical lapses as critically as we now judge Charles Davenport ( Paul, 2014 )? Only time will tell.
Recipient(s) will receive an email with a link to 'The Misuse of Pedigree Analysis in the Eugenics Movement' and will not need an account to access the content.
Subject: The Misuse of Pedigree Analysis in the Eugenics Movement
(Optional message may have a maximum of 1000 characters.)
Citing articles via
Email alerts, affiliations.
- Recent Content
- Browse Issues
- All Content
- Info for Authors
- Info for Librarians
- Editorial Team
- Online ISSN 1938-4211
- Print ISSN 0002-7685
- Copyright © 2024 National Association of Biology Teachers. All rights reserved.
Stay Informed
Disciplines.
- Ancient World
- Anthropology
- Communication
- Criminology & Criminal Justice
- Film & Media Studies
- Food & Wine
- Browse All Disciplines
- Browse All Courses
- Book Authors
- Booksellers
- Instructions
- Journal Authors
- Journal Editors
- Media & Journalists
- Planned Giving
About UC Press
- Press Releases
- Seasonal Catalog
- Acquisitions Editors
- Customer Service
- Exam/Desk Requests
- Media Inquiries
- Print-Disability
- Rights & Permissions
- UC Press Foundation
- © Copyright 2024 by the Regents of the University of California. All rights reserved. Privacy policy Accessibility
This Feature Is Available To Subscribers Only
Sign In or Create an Account
We use cookies to enhance our website for you. Proceed if you agree to this policy or learn more about it.
- Essay Database >
- Essay Examples >
- Essays Topics >
- Essay on Women
Pedigree Case Analysis Essay Sample
Type of paper: Essay
Topic: Women , Gene , Information , Genetics , Parents , Design , Family , Inheritance
Published: 03/12/2020
ORDER PAPER LIKE THIS
Pedigree Case Analysis
A verified pedigree can help in the determination of the most likely pattern of inheritance of a single-gene disorder in a family. Pedigree analysis also has relevance to the determination of approaches to testing, assessment of recurrence risks, and in some instances, prognosis (Leonard, 2007). The purpose of this paper is multi-pronged. It will analyze the pedigree of a family with the aims of identifying the pattern of inheritance of a trait, additional information required, other relatives at risk, and alternative explanations for the condition.
Additional Information Required
The pedigree illustration provided does not depict the presence of the condition in the preceding two generations on both the paternal and maternal side. Further, it does not depict the presence of carriers on either the maternal or paternal side of the family. The additional information required in this case, therefore, is a complete family history for three generations (Cummings, 2013). This information should focus on close relatives on both the paternal and maternal side. It should incorporate the medical histories of grandparents, uncles, aunts, cousins, nephews, and nieces. This history should help elicit information about the presence of the disease or condition in other family generations on each side of the family. A complete family medical history is essential in determining the pattern of inheritance of a condition (Leonard, 2007).
Pattern of Disease Inheritance
Autosomal Dominant Inheritance Based on the limited information available, the likely modes of inheritance of the trait depicted in the pedigree are autosomal dominant or X-linked dominant (Cummings, 2013). Conditions inherited in a dominant fashion are expressed in persons with only one copy of the mutant gene. In an autosomal dominant pattern of inheritance, the mutant allele is located on one of the twenty two pairs of autosomes. Affected males and females have an equal likelyhood of passing the trait to their children due to its being dominant (Westman, 2006). Affected persons have one copy of the normal gene and one copy of the mutant gene. Therefore, every child has a 50% probability of inheriting the mutant gene. This is akin to the pedigree which shows that two of the children of these parents have inherited the condition while the other two have not.
X-linked Dominant Inheritance
In an X-linked inheritance pattern, the gene responsible for a phenotype is located on the X chromosome. Because sons do not inherit the X chromosome from their fathers, there can be no transmission from a father to a son. There can, however, be transmission from a father to a daughter. Regarding the mother, children of an affected female have a 50% probability of inheriting the X chromosome on which the mutant gene is located. Therefore, the trait depicted in the pedigree is likely to have been inherited from the mother in an X-linked dominant fashion. This is because only one daughter and one son have inherited the trait. If the defective trait was located on the father’s X-chromosome, both daughters would have inherited the trait which is not the case (Westman, 2006).
Relatives at Risk
If the condition was inherited in an autosomal dominant fashion, either parent is at risk of the condition. This is because the children must have inherited the defective gene from either parent. The siblings to the affected children are also at risk as it is possible that the condition is yet to manifest itself. If the condition was inherited in an x-linked fashion, the mother is at risk of the condition. This is because the father is unlikely to be the source of the mutant gene since only one daughter is affected.
Alternative Explanations
Alternative explanations for the above condition are the probability that the trait manifesting in the two offspring’s is controlled by more than one gene. Research has shown that the inheritance of human genetic conditions is more complex than previously thought. It involves more than the single gene patterns first described by Mendel. Other patterns of transmission of genetic disorders include mitochondrial inheritance, chromosomal inheritance, and many other atypical patterns of inheritance such as uniparental disomy, imprinting, contiguous gene disorders, and multifactorial inheritance. In addition to the many inheritance patterns, research also shows that the expression of a trait is influenced by environmental factors. Therefore, the trait in the pedigree can be explained by other patterns of inheritance as opposed to the single gene loci pattern and by environmental factors (Leonard, 2007).
In summary, this paper has analyzed the pedigree of a family. It has identified the additional information required in relation to the pedigree. Further, it has identified the likely pattern of inheritance of the condition. In addition, it has identified the relatives at risk of the condition. Lastly, it has described alternative explanations to the condition.
Cummings, M. R. (2013). Human heredity: Principles and issues. Belmont, CA: Cengage Learning. Leonard, D. G. B. (2007). Molecular pathology in clinical practice. New York, NY: Springer Science. Starr, C., Evers, C., & Starr, L. (2011). Biology: Concepts and applications. Belmont, CA: Cengage Learning. Westman, J. A. (2006). Medical genetics for the modern clinician. Baltimore: Lippincott, Williams, & Wilkins.

Cite this page
Share with friends using:
Removal Request

Finished papers: 431
This paper is created by writer with
ID 281578926
If you want your paper to be:
Well-researched, fact-checked, and accurate
Original, fresh, based on current data
Eloquently written and immaculately formatted
275 words = 1 page double-spaced

Get your papers done by pros!
Other Pages
Domestic violence annotated bibliographies, graphic term papers, manifestation term papers, greatness term papers, array term papers, gaze term papers, butte essays, convertible bond essays, assent essays, ascariasis essays, federal reserve board essays, lobes essays, nitrogen balance essays, ready made essays, ejaculatory essays, freshman year essays, gonyea essays, hostels essays, saudi gazette essays, philo essays, public figures essays, champagne essays, sleeves essays, zurich essays, robbers essays, capital account essays, political me essay examples, the women s army corp research paper example, comparing and contrasting two articles essay examples, american history great depression 20th century the gilded age and world war essay sample, example of essay on ethics in the near future, essay on independent living, example of report on what is greece s current economic situation, aviation and regulatory agencies book review examples, free case study about coca cola share a coke campaign, free performing arts and gender constitution article review example, example of chain restaurant project report, good example of clifton metal works mgmt case study, free literature review on even in the midst of his panic winston was too much taken aback to be able to hold his tongue, free research proposal on the effects of rmb hkd exchange rate on the hong kong retail market, nursing theory essay sample, marketing ethics essay samples.
Password recovery email has been sent to [email protected]
Use your new password to log in
You are not register!
By clicking Register, you agree to our Terms of Service and that you have read our Privacy Policy .
Now you can download documents directly to your device!
Check your email! An email with your password has already been sent to you! Now you can download documents directly to your device.
or Use the QR code to Save this Paper to Your Phone
The sample is NOT original!
Short on a deadline?
Don't waste time. Get help with 11% off using code - GETWOWED
No, thanks! I'm fine with missing my deadline
Advertisement
Supported by
A Climate Scientist Is Voted President of an Oil Country. Now What?
Claudia Sheinbaum, Mexico’s next leader, is an academic and a politician. Here’s what her track record reveals.
- Share full article

By Somini Sengupta
Mexico is the world’s 11th-largest oil producer. It has been gripped by a deadly heat wave. Now, it’s elected as its president a woman with a rare pedigree: a left-of-center climate scientist with a doctorate in energy engineering named Claudia Sheinbaum.
Ms. Sheinbaum is no stranger to politics nor to environmental crises. She was mayor of Mexico City, a vibrant metropolitan area of 23 million that faces a dire water crisis. She helped write the Intergovernmental Panel on Climate Change reports, the sweeping United Nations documents that have warned the world about the hazards of burning fossil fuels.
Ms. Sheinbaum will have to balance numerous, sometimes contradictory, tests as she takes office. Federal budgets are tight. Energy demands are rising. Mexico’s national oil company is heavily indebted. She’ll face the challenges of poverty, migration, organized crime and relations with the next president of the United States.
It would be folly to predict what she will do, but it’s worth looking at what she has said and done on energy and environmental issues so far in her career.
First, her record.
As mayor of Mexico City, she began electrifying the city’s public bus fleet. She set up a huge rooftop solar array on the city’s main wholesale market. She expanded bike lanes, making permanent several kilometers of pandemic-era pop-up paths.
She has been criticized by environmentalists for backing one of the country’s most controversial infrastructure projects, the 1,500-kilometer so-called Maya Train corridor, which cuts across forests and archaeological sites to connect tourist sites like Cancún to rural areas on the Yucatán Peninsula.
We are having trouble retrieving the article content.
Please enable JavaScript in your browser settings.
Thank you for your patience while we verify access. If you are in Reader mode please exit and log into your Times account, or subscribe for all of The Times.
Thank you for your patience while we verify access.
Already a subscriber? Log in .
Want all of The Times? Subscribe .
clock This article was published more than 1 year ago
We tested a new ChatGPT-detector for teachers. It flagged an innocent student.
Five high school students helped our tech columnist test a ChatGPT detector coming from Turnitin to 2.1 million teachers. It missed enough to get someone in trouble.

High school senior Lucy Goetz got the highest possible grade on an original essay she wrote about socialism. So imagine her surprise when I told her that a new kind of educational software I’ve been testing claimed she got help from artificial intelligence.
A new AI-writing detector from Turnitin — whose software is already used by 2.1 million teachers to spot plagiarism — flagged the end of her essay as likely being generated by ChatGPT .
“Say what?” says Goetz, who swears she didn’t use the AI writing tool to cheat. “I’m glad I have good relationships with my teachers.”
After months of sounding the alarm about students using AI apps that can churn out essays and assignments, teachers are getting AI technology of their own. On April 4, Turnitin is activating the software I tested for some 10,700 secondary and higher-educational institutions, assigning “generated by AI” scores and sentence-by-sentence analysis to student work. It joins a handful of other free detectors already online. For many teachers I’ve been hearing from, AI detection offers a weapon to deter a 21st-century form of cheating.
But AI alone won’t solve the problem AI created. The flag on a portion of Goetz’s essay was an outlier, but shows detectors can sometimes get it wrong — with potentially disastrous consequences for students. Detectors are being introduced before they’ve been widely vetted, yet AI tech is moving so fast, any tool is likely already out of date.
It’s a pivotal moment for educators: Ignore AI and cheating could go rampant. Yet even Turnitin’s executives tell me that treating AI purely as the enemy of education makes about as much sense in the long run as trying to ban calculators.
Ahead of Turnitin’s launch this week, the company says 2 percent of customers have asked it not to display the AI writing score on student work. That includes a "significant majority” of universities in the United Kingdom, according to UCISA , a professional body for digital educators.
To see what’s at stake, I asked Turnitin for early access to its software. Five high school students, including Goetz, volunteered to help me test it by creating 16 samples of real, AI-fabricated and mixed-source essays to run past Turnitin’s detector.
The result? It got over half of them at least partly wrong. Turnitin accurately identified six of the 16 — but failed on three, including a flag on 8 percent of Goetz’s original essay. And I’d give it only partial credit on the remaining seven, where it was directionally correct but misidentified some portion of ChatGPT-generated or mixed-source writing.
Turnitin claims its detector is 98 percent accurate overall. And it says situations such as what happened with Goetz’s essay, known as a false positive, happen less than 1 percent of the time, according to its own tests.
Turnitin also says its scores should be treated as an indication, not an accusation . Still, will millions of teachers understand they should treat AI scores as anything other than fact? After my conversations with the company, it added a caution flag to its score that reads, “Percentage may not indicate cheating. Review required.”
“Our job is to create directionally correct information for the teacher to prompt a conversation,” Turnitin chief product officer Annie Chechitelli tells me. “I’m confident enough to put it out in the market, as long as we’re continuing to educate educators on how to use the data.” She says the company will keep adjusting its software based on feedback and new AI advancements.
The question is whether that will be enough. “The fact that the Turnitin system for flagging AI text doesn’t work all the time is concerning,” says Rebecca Dell, who teaches Goetz’s AP English class in Concord, Calif. “I’m not sure how schools will be able to definitively use the checker as ‘evidence’ of students using unoriginal work.”
Unlike accusations of plagiarism, AI cheating has no source document to reference as proof. “This leaves the door open for teacher bias to creep in,” says Dell.
For students, that makes the prospect of being accused of AI cheating especially scary. “There is no way to prove that you didn’t cheat unless your teacher knows your writing style, or trusts you as a student,” says Goetz.
Why detecting AI is so hard
Spotting AI writing sounds deceptively simple. When a colleague recently asked me if I could detect the difference between real and ChatGPT-generated emails, I didn’t perform very well.
Detecting AI writing with software involves statistics. And statistically speaking, the thing that makes AI distinct from humans is that it’s “extremely consistently average,” says Eric Wang, Turnitin’s vice president of AI.
Systems such as ChatGPT work like a sophisticated version of auto-complete, looking for the most probable word to write next. “That’s actually the reason why it reads so naturally: AI writing is the most probable subset of human writing,” he says.
Turnitin’s detector “identifies when writing is too consistently average,” Wang says.
The challenge is that sometimes a human writer may actually look consistently average.
On economics, math and lab reports, students tend to hew to set styles, meaning they’re more likely to be misidentified as AI writing, says Wang. That’s likely why Turnitin erroneously flagged Goetz’s essay, which veered into economics. (“My teachers have always been fairly impressed with my writing,” says Goetz.)
Wang says Turnitin worked to tune its systems to err on the side of requiring higher confidence before flagging a sentence as AI. I saw that develop in real time: I first tested Goetz’s essay in late January, and the software identified much more of it — about 50 percent — as being AI generated. Turnitin ran my samples through its system again in late March, and that time only flagged 8 percent of Goetz’s essay as AI-generated.
But tightening up the software’s tolerance came with a cost: Across the second test of my samples, Turnitin missed more actual AI writing. “We’re really emphasizing student safety,” says Chechitelli.
Say hello to your new tutor: It’s ChatGPT
Turnitin does perform better than other public AI detectors I tested. One introduced in February by OpenAI, the company that invented ChatGPT, got eight of our 16 test samples wrong. (Independent tests of other detectors have declared they “ fail spectacularly .”)
Turnitin’s detector faces other important technical limitations, too. In the six samples it got completely right, they were all clearly 100 percent student work or produced by ChatGPT. But when I tested it with essays from mixed AI and human sources, it often misidentified the individual sentences or missed the human part entirely. And it couldn’t spot the ChatGPT in papers we ran through Quillbot, a paraphrasing program that remixes sentences.
What’s more, Turnitin’s detector may already be behind the state of the AI art. My student helpers created samples with ChatGPT, but since they did the writing, the app has gotten a software update called GPT-4 with more creative and stylistic capabilities. Google also introduced a new AI bot called Bard . Wang says addressing them is on his road map.
Some AI experts say any detection efforts are at best setting up an arms race between cheaters and detectors. “I don’t think a detector is long-term reliable,” says Jim Fan, an AI scientist at Nvidia who used to work at OpenAI and Google.
“The AI will get better, and will write in ways more and more like humans. It is pretty safe to say that all of these little quirks of language models will be reduced over time,” he says.
Is detecting AI a good idea?
Given the potential — even at 1 percent — of being wrong, why release an AI detector into software that will touch so many students?
“Teachers want deterrence,” says Chechitelli. They’re extremely worried about AI and helping them see the scale of the actual problem will “bring down the temperature.”
Some educators worry it will actually raise the temperature.
Mitchel Sollenberger, the associate provost for digital education at the University of Michigan-Dearborn, is among the officials who asked Turnitin not to activate AI detection for his campus at its initial launch.
He has specific concerns about how false positives on the roughly 20,000 student papers his faculty run through Turnitin each semester could lead to baseless academic-integrity investigations. “Faculty shouldn’t have to be expert in a third-party software system — they shouldn’t necessarily have to understand every nuance,” he says.
Ian Linkletter, who serves as emerging technology and open-education librarian at the British Columbia Institute of Technology, says the push for AI detectors reminds him of the debate about AI exam proctoring during pandemic virtual learning.
“I am worried they’re marketing it as a precision product, but they’re using dodgy language about how it shouldn’t be used to make decisions,” he says. “They’re working at an accelerated pace not because there is any desperation to get the product out but because they’re terrified their existing product is becoming obsolete.”
Said Chechitelli: “We are committed to transparency with the community and have been clear about the need to continue iterating on the user experience as we learn more from students and educators.
Deborah Green, CEO of UCISA in the U.K., tells me she understands and appreciates Turnitin’s motives for the detector. “What we need is time to satisfy ourselves as to the accuracy, the reliability and particularly the suitability of any tool of this nature.”
It’s not clear how the idea of an AI detector fits into where AI is headed in education . “In some academic disciplines, AI tools are already being used in the classroom and in assessment,” says Green. “The emerging view in many U.K. universities is that with AI already being used in many professions and areas of business, students actually need to develop the critical thinking skills and competencies to use and apply AI well.”
There’s a lot more subtlety to how students might use AI than a detector can flag today.
My student tests included a sample of an original student essay written in Spanish, then translated into English with ChatGPT. In that case, what should count: the ideas or the words? What if the student was struggling with English as a second language? (In our test, Turnitin’s detector appeared to miss the AI writing, and flagged none of it.)
Would it be more or less acceptable if a student asked ChatGPT to outline all the ideas for an assignment, and then wrote the actual words themselves?
“That’s the most interesting and most important conversation to be having in the next six months to a year — and one we’ve been having with instructors ourselves,” says Chechitelli.
“We really feel strongly that visibility, transparency and integrity are the foundations of the conversations we want to have next around how this technology is going to be used,” says Wang.
For Dell, the California teacher, the foundation of AI in the classroom is an open conversation with her students.
When ChatGPT first started making headlines in December, Dell focused an entire lesson with Goetz’s English class on what ChatGPT is, and isn’t good for. She asked it to write an essay for an English prompt her students had already completed themselves, and then the class analyzed the AI’s performance.
The AI wasn’t very good.
“Part of convincing kids not to cheat is making them understand what we ask them to do is important for them,” said Dell.
Help Desk: Making tech work for you
Help Desk is a destination built for readers looking to better understand and take control of the technology used in everyday life.
Take control: Sign up for The Tech Friend newsletter to get straight talk and advice on how to make your tech a force for good.
Tech tips to make your life easier: 10 tips and tricks to customize iOS 16 | 5 tips to make your gadget batteries last longer | How to get back control of a hacked social media account | How to avoid falling for and spreading misinformation online
Data and Privacy: A guide to every privacy setting you should change now . We have gone through the settings for the most popular (and problematic) services to give you recommendations. Google | Amazon | Facebook | Venmo | Apple | Android
Ask a question: Send the Help Desk your personal technology questions .

IMAGES
VIDEO
COMMENTS
Pedigree analysis was developed to understand the inheritance of genes from parents to offspring. It was developed as a chart that can represent a family tree along with the family members and their genetic traits, respectively. Gregor Mendel's experiments showed that the "factors" that we now know as "genes", are the factors which ...
This page titled 4: Pedigree Analysis is shared under a CC BY-NC-SA 4.0 license and was authored, remixed, and/or curated by Natasha Ramroop Singh via source content that was edited to the style and standards of the LibreTexts platform; a detailed edit history is available upon request.
A pedigree analysis chart provides an important tool to study the inheritance of genes. It also gives precise information about the family members through diagrammatic representation. Sometimes, it is also known as the " family or genealogic tree ". A pedigree chart uses internationally accepted symbols, lines, numbers and letters To ...
Pedigree charts are used by scientists, genetic counselors, and animal breeders. Pedigrees show inheritance across one or more generations. Individuals connected by a horizontal line have mated and had children. Vertical lines connect parents to their children. Siblings are generally shown from left to right according to birth order.
Matings are drawn as a line joining a male and female, while a consanguineous mating (closely related is two lines. Figure 5.2.2 5.2. 2: Symbols used in drawing a pedigree. (Original-Deyholos-CC:AN) The affected individual that brings the family to the attention of a geneticist is called the proband (or propositus).
Pedigree. Chart that shows the presence or absence of a trait within a family across generations. Genotype. The genetic makeup of an organism (ex: TT) Phenotype. The physical characteristics of an organism (ex: tall) Dominant allele. Allele that is phenotypically expressed over another allele. Recessive allele.
This consists of three stages: Organizing the facts that you have. Estimating dates and places. Planning the next research step. 1. Organizing the Facts That You Have. The following chart shows part of my pedigree with the known facts organized in standard fashion. Pedigree Analysis: Step 1-Organizing Known Facts. 2.
Therefore, Pedigree analysis is an important tool in basic research and genetic counseling. Each pedigree chart represents all available information about the inheritance of a single trait (most often a disease) within a family. The pedigree chart is therefore drawn using factual information. Still, there is always some possibility of errors in ...
An approach, called pedigree analysis, is used to study the inheritance of genes in humans. This session will outline how to construct a family pedigree, and how to interpret the information in a pedigree using Mendel's laws of inheritance and an understanding of the chromosome theory of inheritance. To construct a pedigree based on a family ...
Pedigree Analysis (cf. chapters 4.4, 5.2, 6.2 of textbook) • "crosses" and "matings" in human(s) (families) are shown and analyzed in pedigrees - pedigrees are formalized ways using standard sets of symbols to depict family trees and lineages - pedigrees provide concise and accurate records of families
Pedigree Analysis Dr. Monisha Banerjee Professor Molecular & Human Genetics Lab Department of Zoology University of Lucknow Lucknow A pedigree is a pictorial representation of a family history. It is an important tool for studying inherited diseases and other traits. ...
Pedigree Analysis In genetics, pedigree analysis or family history is a method of determining the probability of a genetic disease being inherited from one's parents. This probability is expressed as a simple ratio of the number of affected relatives to the total number of relatives. The most common form of pedigree analysis is used to ...
Types of Pedigree Analysis. Based on linked chromosomes, pedigree analysis is classified into: 1) Sex-linked pedigree. It is subdivided into: X-linked pedigree. Y-linked pedigree. Even the X-linked pedigree is further divided into: X-linked dominant pedigree. X-linked recessive pedigree.
Pedigree analysis is a self-study assignment. As a component of their directed study assignment, students are asked to work out multiple pedigree problems individually. After a few days, students have the opportunity to attend a Q&A session with the instructor. During this session, students are encouraged to ask the instructor specific ...
Definition of Pedigree Analysis in Biology. Pedigree analysis is an examination, or demonstration of the inheritance pattern of particular trait(s) in human beings. It can be represented by a pedigree chart, which is a visual representation of a family tree linking family members and their genetic trail.. In the context of genetics, medicine, and biology, t hese traits are typically diseases ...
Healthcare professionals have long known that common diseases (e.g., heart disease, cancer, and diabetes) and rare diseases (e.g., hemophilia, cystic fibrosis, and sickle cell anemia) can run in families. For example, if one generation of a family has high blood pressure, it is not unusual for the next generation to have similarly high blood pressure. Family history can be a powerful screening ...
The analysis of pedigree problems is time-consuming, and all results described here were derived from the analysis of only one pedigree per type of autosomal inheritance. Based on the available data and results, it is therefore not possible to make any predictions about the general strategic approach of secondary students to other pedigrees ...
The Family Pedigree Analysis, Research Paper Example. HIRE A WRITER! You are free to use it as an inspiration or a source for your own work. After carefully analyzing my family pedigree, there are two important trends to notice with regards to diseases passed along throughout the family. The first disease of note is on my parental side by which ...
Pedigree analysis has long been an essential tool in human genetics as well as a staple of genetics education. Students of genetics might be surprised to learn that human pedigrees were first popularized in the United States by proponents of eugenics, the pseudoscientific social movement aimed at improving the genetic quality of the human race. Notably, the influential eugenicist Charles B ...
A verified pedigree can help in the determination of the most likely pattern of inheritance of a single-gene disorder in a family. Pedigree analysis also has relevance to the determination of approaches to testing, assessment of recurrence risks, and in some instances, prognosis (Leonard, 2007). The purpose of this paper is multi-pronged.
Accepted: 20 June 2017/Published online: 28 June 2017 The Author(s) 2017. This article is an open access publication. Abstract Pedigree dog breeding has been the subject of public debate due to health problems caused by breeding for extreme looks and the narrow genepool of many breeds. Our research aims to provide insights in order to further ...
Harvard College Writing Center 5 Asking Analytical Questions When you write an essay for a course you are taking, you are being asked not only to create a product (the essay) but, more importantly, to go through a process of thinking more deeply about a question or problem related to the course. By writing about a
Leer en español. Mexico is the world's 11th-largest oil producer. It has been gripped by a deadly heat wave. Now, it's elected as its president a woman with a rare pedigree: a left-of-center ...
It flagged an innocent student. Five high school students helped our tech columnist test a ChatGPT detector coming from Turnitin to 2.1 million teachers. It missed enough to get someone in trouble ...
Tips for writing a successful MBA statement of purpose. As you write your SOP, here are a few things to keep in mind that can help your writing stand out: Clearly state your goals: Openly communicate your short-term and long-term goals in earning your MBA. Clear statements around this crucial element of your SOP can help you avoid any potential ...
Artificial intelligence (AI) is the theory and development of computer systems capable of performing tasks that historically required human intelligence, such as recognizing speech, making decisions, and identifying patterns. AI is an umbrella term that encompasses a wide variety of technologies, including machine learning, deep learning, and ...