February 16, 2016

What Sparked the Cambrian Explosion?
An evolutionary burst 540 million years ago filled the seas with an astonishing diversity of animals. The trigger behind that revolution is finally coming into focus
By Douglas Fox & Nature magazine

Given the importance of oxygen for animals, researchers suspected that a sudden increase in the gas to near-modern levels in the ocean could have spurred the Cambrian explosion. To test that idea, they have studied ancient ocean sediments laid down during the Ediacaran and Cambrian periods, which together ran from about 635 million to 485 million years ago.
©iStock
A series of dark, craggy pinnacles rises 80 meters above the grassy plains of Namibia. The peaks call to mind something ancient — the burial mounds of past civilizations or the tips of vast pyramids buried by the ages.
The stone formations are indeed monuments of a faded empire, but not from anything hewn by human hands. They are pinnacle reefs, built by cyanobacteria on the shallow sea floor 543 million years ago, during a time known as the Ediacaran period. The ancient world occupied by these reefs was truly alien. The oceans held so little oxygen that modern fish would quickly founder and die there. A gooey mat of microbes covered the sea floor at the time, and on that blanket lived a variety of enigmatic animals whose bodies resembled thin, quilted pillows. Most were stationary, but a few meandered blindly over the slime, grazing on the microbes. Animal life at this point was simple, and there were no predators. But an evolutionary storm would soon upend this quiet world.
Within several million years, this simple ecosystem would disappear, and give way to a world ruled by highly mobile animals that sported modern anatomical features. The Cambrian explosion , as it is called, produced arthropods with legs and compound eyes, worms with feathery gills and swift predators that could crush prey in tooth-rimmed jaws. Biologists have argued for decades over what ignited this evolutionary burst. Some think that a steep rise in oxygen sparked the change, whereas others say that it sprang from the development of some key evolutionary innovation, such as vision. The precise cause has remained elusive, in part because so little is known about the physical and chemical environment at that time.
On supporting science journalism
If you're enjoying this article, consider supporting our award-winning journalism by subscribing . By purchasing a subscription you are helping to ensure the future of impactful stories about the discoveries and ideas shaping our world today.
But over the past several years, discoveries have begun to yield some tantalizing clues about the end of the Ediacaran. Evidence gathered from the Namibian reefs and other sites suggests that earlier theories were overly simplistic — that the Cambrian explosion actually emerged out of a complex interplay between small environmental changes that triggered major evolutionary developments.
Some scientists now think that a small, perhaps temporary, increase in oxygen suddenly crossed an ecological threshold, enabling the emergence of predators. The rise of carnivory would have set off an evolutionary arms race that led to the burst of complex body types and behaviours that fill the oceans today. “This is the most significant event in Earth evolution,” says Guy Narbonne, a palaeobiologist at Queen's University in Kingston, Canada. “The advent of pervasive carnivory, made possible by oxygenation, is likely to have been a major trigger.”
Energy to burn
In the modern world, it's easy to forget that complex animals are relative newcomers to Earth. Since life first emerged more than 3 billion years ago, single-celled organisms have dominated the planet for most of its history. Thriving in environments that lacked oxygen, they relied on compounds such as carbon dioxide, sulfur-containing molecules or iron minerals that act as oxidizing agents to break down food. Much of Earth's microbial biosphere still survives on these anaerobic pathways.
Animals, however, depend on oxygen — a much richer way to make a living. The process of metabolizing food in the presence of oxygen releases much more energy than most anaerobic pathways. Animals rely on this potent, controlled combustion to drive such energy-hungry innovations as muscles, nervous systems and the tools of defence and carnivory — mineralized shells, exoskeletons and teeth.
In Namibia, China and other spots around the world, researchers have collected rocks that were once ancient seabeds, and analysed the amounts of iron, molybdenum and other metals in them. The metals' solubility depends strongly on the amount of oxygen present, so the amount and type of those metals in ancient sedimentary rocks reflect how much oxygen was in the water long ago, when the sediments formed.
These proxies seemed to indicate that oxygen concentrations in the oceans rose in several steps, approaching today's sea-surface concentrations at the start of the Cambrian, around 541 million years ago — just before more-modern animals suddenly appeared and diversified. This supported the idea of oxygen as a key trigger for the evolutionary explosion.
But last year, a major study of ancient sea-floor sediments challenged that view. Erik Sperling, a palaeontologist at Stanford University in California, compiled a database of 4,700 iron measurements taken from rocks around the world, spanning the Ediacaran and Cambrian periods. He and his colleagues did not find a statistically significant increase in the proportion of oxic to anoxic water at the boundary between the Ediacaran and the Cambrian.
“Any oxygenation event must have been far, far smaller than what people normally considered,” concludes Sperling. Most people assume “that the oxygenation event essentially raised oxygen to essentially modern-day levels. And that probably wasn't the case”, he says.
The latest results come at a time when scientists are already reconsidering what was happening to ocean oxygen levels during this crucial period. Donald Canfield, a geobiologist at the University of Southern Denmark in Odense, doubts that oxygen was a limiting factor for early animals. In a study published last month, he and his colleagues suggest that oxygen levels were already high enough to support simple animals, such as sponges, hundreds of millions of years before they actually appeared. Cambrian animals would have needed more oxygen than early sponges, concedes Canfield. “But you don't need an increase in oxygen across the Ediacaran/Cambrian boundary,” he says; oxygen could already have been abundant enough “for a long, long time before”.
“The role of oxygen in the origins of animals has been heavily debated,” says Timothy Lyons, a geobiologist at the University of California, Riverside. “In fact, it's never been more debated than it is now.” Lyons sees a role for oxygen in evolutionary changes, but his own work with molybdenum and other trace metals suggests that the increases in oxygen just before the Cambrian were mostly temporary peaks that lasted a few million years and gradually stepped upward (see 'When life sped up' ).
Modern mirrors
Sperling has looked for insights into Ediacaran oceans by studying oxygen-depleted regions in modern seas around the globe. He suggests that biologists have conventionally taken the wrong approach to thinking about how oxygen shaped animal evolution. By pooling and analysing previously published data with some of his own, he found that tiny worms survive in areas of the sea floor where oxygen levels are incredibly low — less than 0.5% of average global sea-surface concentrations. Food webs in these oxygen-poor environments are simple, and the animals feed directly on microbes. In places where sea-floor oxygen levels are a bit higher — about 0.5–3% of concentrations at the sea surface — animals are more abundant but their food webs remain limited: the animals still feed on microbes rather than on each other. But around somewhere between 3% and 10% oxygen levels, predators emerge and start to consume other animals.
The implications of this finding for evolution are profound, Sperling says.The modest oxygen rise that he thinks may have occurred just before the Cambrian would have been enough to trigger a big change. “If oxygen levels were 3% and they rose past that 10% threshold, that would have had a huge influence on early animal evolution,” he says. “There's just so much in animal ecology, lifestyle and body size that seems to change so dramatically through those levels.”
The gradual emergence of predators, driven by a small rise in oxygen, would have meant trouble for Ediacaran animals that lacked obvious defences. “You're looking at soft-bodied, mostly immobile forms that probably lived their lives by absorbing nutrients through their skin,” says Narbonne.
Studies of those ancient Namibian reefs suggest that animals were indeed starting to fall prey to predators by the end of the Ediacaran. When palaeobiologist Rachel Wood from the University of Edinburgh, UK, examined the rock formations, she found spots where a primitive animal called C loudina had taken over parts of the microbial reef. Rather than spreading out over the ocean floor, these cone-shaped creatures lived in crowded colonies, which hid their vulnerable body parts from predators — an ecological dynamic that occurs in modern reefs.
C loudina were among the earliest animals known to have grown hard, mineralized exoskeletons. But they were not alone. Two other types of animal in those reefs also had mineralized parts, which suggests that multiple, unrelated groups evolved skeletal shells around the same time. “Skeletons are quite costly to produce,” says Wood. “It's very difficult to come up with a reason other than defence for why an animal should bother to create a skeleton for itself.” Wood thinks that the skeletons provided protection against newly evolved predators. Some C loudina fossils from that period even have holes in their sides, which scientists interpret as the marks of attackers that bore into the creatures' shells.
Palaeontologists have found other hints that animals had begun to eat each other by the late Ediacaran. In Namibia, Australia and Newfoundland in Canada, some sea-floor sediments have preserved an unusual type of tunnel made by an unknown, wormlike creature. Called Treptichnus burrows, these warrens branch again and again, as if a predator just below the microbial mat had systematically probed for prey animals on top. The Treptichnus burrows resemble those of modern priapulid, or 'penis', worms — voracious predators that hunt in a remarkably similar way on modern sea floors.
The rise of predation at this time put large, sedentary Ediacaran animals at a big disadvantage. “Sitting around doing nothing becomes a liability,” says Narbonne.
The world in 3D
The moment of transition from the Ediacaran to the Cambrian world is recorded in a series of stone outcrops rounded by ancient glaciers on the south edge of Newfoundland. Below that boundary are impressions left by quilted Ediacaran animals, the last such fossils recorded on Earth. And just 1.2 meters above them, the grey siltstone holds trails of scratch marks, thought to have been made by animals with exoskeletons, walking on jointed legs — the earliest evidence of arthropods in Earth's history.
No one knows how much time passed in that intervening rock — maybe as little as a few centuries or millennia, says Narbonne. But during that short span, the soft-bodied, stationary Ediacaran fauna suddenly disappeared, driven to extinction by predators, he suggests.
Narbonne has closely studied the few fauna that survived this transition, and his findings suggest that some of them had acquired new, more complex types of behaviour. The best clues come from traces left by peaceful, wormlike animals that grazed on the microbial mat. Early trails from about 555 million years ago meander and criss-cross haphazardly, indicating a poorly developed nervous system that was unable to sense or react to other grazers nearby — let alone predators. But at the end of the Ediacaran and into the early Cambrian, the trails become more sophisticated: creatures carved tighter turns and ploughed closely spaced, parallel lines through the sediments. In some cases, a curvy feeding trail abruptly transitions into a straight line, which Narbonne interprets as potential evidence of the grazer evading a predator.
This change in grazing style may have contributed to the fragmentation of the microbial mat, which began early in the Cambrian. And the transformation of the sea floor, says Narbonne, “may have been the most profound change in the history of life on Earth”, . The mat had previously covered the seabed like a coating of plastic wrap, leaving the underlying sediments largely anoxic and off limits to animals. Because animals could not burrow deeply in the Ediacaran, he says, “the mat meant that life was two-dimensional”. When grazing capabilities improved, animals penetrated the mat and made the sediments habitable for the first time, which opened up a 3D world.
Tracks from the early Cambrian show that animals started to burrow several centimeters into the sediments beneath the mat, which provided access to previously untapped nutrients — as well as a refuge from predators. It's also possible that animals went in the opposite direction. Sperling says that the need to avoid predators (and pursue prey) may have driven animals into the water column above the seabed, where enhanced oxygen levels enabled them to expend energy through swimming.
The emerging evidence about oxygen thresholds and ecology could also shed light on another major evolutionary question: when did animals originate? The first undisputed fossils of animals appear only 580 million years ago, but genetic evidence indicates that basic animal groups originated as far back as 700 million to 800 million years ago. According to Lyons, the solution may be that oxygen levels rose to perhaps 2% or 3% of modern levels around 800 million years ago. These concentrations could have sustained small, simple animals, just as they do today in the ocean's oxygen-poor zones. But animals with large bodies could not have evolved until oxygen levels climbed higher in the Ediacaran.
Understanding how oxygen influenced the appearance of complex animals will require scientists to tease more-subtle clues out of the rocks. “We've been challenging people working on fossils to tie their fossils more closely to our oxygen proxies,” says Lyons. It will mean deciphering what oxygen levels were in different ancient environments, and connecting those values with the kinds of traits exhibited by the animal fossils found in the same locations.
This past autumn, Woods visited Siberia with that goal in mind. She collected fossils of Cloudina and another skeletonized animal, Suvorovella , from the waning days of the Ediacaran. Those sites gave her the chance to gather fossils from many different depths in the ancient ocean, from the more oxygen-rich surface waters to deeper zones. Wood plans to look for patterns in where animals were growing tougher skeletons, whether they were under attack by predators and whether any of this had a clear link with oxygen levels, she says. “Only then can you pick out the story.”
This article is reproduced with permission and was first published on February 16, 2016.
Current understanding on the Cambrian Explosion: questions and answers
- Paläontologische Zeitschrift 95(4)

- Northwest University

Abstract and Figures

Discover the world's research
- 25+ million members
- 160+ million publication pages
- 2.3+ billion citations

- GONDWANA RES

- PALAEOGEOGR PALAEOCL

- Alexander J. Dickson

- Volker Thiel

- EARTH-SCI REV

- Stefan Bengtson
- Dian J. Teigler
- Kenneth M. Towe
- Preston E. Cloud

- Xian‐guang Hou
- David J. Siveter
- Derek J. Siveter

- Noah J. Planavsky

- Recruit researchers
- Join for free
- Login Email Tip: Most researchers use their institutional email address as their ResearchGate login Password Forgot password? Keep me logged in Log in or Continue with Google Welcome back! Please log in. Email · Hint Tip: Most researchers use their institutional email address as their ResearchGate login Password Forgot password? Keep me logged in Log in or Continue with Google No account? Sign up
Testing the Cambrian explosion hypothesis by using a molecular dating technique
Information & authors, metrics & citations, view options, sign up for pnas alerts..
Get alerts for new articles, or get an alert when an article is cited.

ABBREVIATION
Acknowledgments, information, published in, classifications.
- Biological Sciences
Submission history
Affiliations, citation statements.
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
View options
Download this article as a PDF file
Login options
Check if you have access through your login credentials or your institution to get full access on this article.
Recommend to a librarian
Purchase options.
Purchase this article to access the full text.
Single Article Purchase
- pp. 12073-12734
Restore content access
Restore content access for purchases made as a guest
Share article link
Copying failed.
Share on social media
Further reading in this issue, supplementary material can now appear online.
- Nicholas R. Cozzarelli
A role for the β-amyloid precursor protein in memory?
- Sangram S. Sisodia ,
- Michela Gallagher and
Cell death throes
- Suzanne Cory and
Physician–patient racial concordance and newborn mortality
- George J. Borjas ,
- Robert VerBruggen ,
Bodily maps of emotions
- Lauri Nummenmaa ,
- Enrico Glerean ,
- Riitta Hari ,
- Jari K. Hietanen ,
Comparing crime rates between undocumented immigrants, legal immigrants, and native-born US citizens in Texas
- Michael T. Light ,
- Jingying He ,
- Jason P. Robey and
Sign up for the PNAS Highlights newsletter
Request username.
Can't sign in? Forgot your username? Enter your email address below and we will send you your username
If the address matches an existing account you will receive an email with instructions to retrieve your username
Create a new account
Change password.
Your Phone has been verified
- History & Society
- Science & Tech
- Biographies
- Animals & Nature
- Geography & Travel
- Arts & Culture
- Games & Quizzes
- On This Day
- One Good Fact
- New Articles
- Lifestyles & Social Issues
- Philosophy & Religion
- Politics, Law & Government
- World History
- Health & Medicine
- Browse Biographies
- Birds, Reptiles & Other Vertebrates
- Bugs, Mollusks & Other Invertebrates
- Environment
- Fossils & Geologic Time
- Entertainment & Pop Culture
- Sports & Recreation
- Visual Arts
- Demystified
- Image Galleries
- Infographics
- Top Questions
- Britannica Kids
- Saving Earth
- Space Next 50
- Student Center

- Why is biology important?

Cambrian explosion
Our editors will review what you’ve submitted and determine whether to revise the article.
- Weizmann Institute of Science - Davidson Institute of Science Education - The Cambrian - An Explosion of Life
- Natural History Museum - The Cambrian explosion was far shorter than we thought
- National Center for Biotechnology Information - PubMed Central - The two phases of the Cambrian Explosion
- Academia - The Cambrian Explosion: macroevolution and biomineralization
- The Geological Society of America - It’s Time to Defuse the Cambrian “Explosion”
- PBS - Evolution - The Cambrian Explosion
- Untamed Science - History of Life: The Cambrian Explosion
- Biology LibreTexts - The Cambrian Explosion
- Nature - What sparked the Cambrian explosion?
Cambrian explosion , the unparalleled emergence of organisms between 541 million and approximately 530 million years ago at the beginning of the Cambrian Period . The event was characterized by the appearance of many of the major phyla (between 20 and 35) that make up modern animal life. Many other phyla also evolved during this time, the great majority of which became extinct during the following 50 to 100 million years. Ironically, many of the most successful modern phyla (including the chordates , which encompass all vertebrates ) are rare elements in Cambrian assemblages; phyla that include the arthropods and sponges contained the most numerically dominant taxa (taxonomic groups) during the Cambrian, and those were the taxa that became extinct.
The beginning of the Cambrian Period is marked by the evolution of hard body parts such as calcium carbonate shells. These body parts fossilize more easily than soft tissues, and thus the fossil record becomes much more complete after their appearance. Many lineages of animals independently evolved hard parts at about the same time. The reasons for this are still debated, but a leading theory is that the amount of oxygen in the atmosphere had finally reached levels that allowed large, complex animals to exist. Oxygen levels may also have facilitated the metabolic processes that produce collagen , a protein building block that is the basis for hard structures in the body.
Other major changes that occurred in the Early Cambrian (541 to 510 million years ago) include the development of animal species that burrowed into the sediments of the seafloor, rather than lying on top of it, and the evolution of the first carbonate reefs, which were built by spongelike animals called archaeocyathids .

By the Early Cambrian the bulk of the biosphere was confined to the margins of the world’s oceans ; no life was found on land (except possibly cyanobacteria [formerly known as blue-green algae ] in moist sediment), relatively few pelagic species (biota living in the open sea) existed, and no organisms inhabited the ocean depths. Life in the shallow regions of the seafloor, however, was already well diversified. This early aquatic ecosystem included the relatively large carnivore Anomalocaris , the deposit-feeding trilobites (early arthropods) and mollusks , the suspension-feeding sponges, various scavenging arthropods, and possibly even parasites such as the onychophoran Aysheaia . Thus, it seems likely that a well-developed aquatic ecosystem was already in operation in the ocean shallows by this time.
Following the Cambrian Period, the biosphere continued to expand relatively rapidly. In the Ordovician Period (485.4 million to 443.4 million years ago), the classic Paleozoic marine faunas—which included bryozoans , brachiopods , corals , nautiloids, and crinoids —developed. Many marine species died off near the end of the Ordovician because of environmental changes. The Silurian Period (443.4 million to 419.2 million years ago) marks a time when a rapid evolution of many suspension-feeders in the oceans occurred. As a result, pelagic predators such as nautiloids became abundant . Gnathostome fishes , the oldest craniates, became common near the end of Silurian times.
Login to your account
If you don't remember your password, you can reset it by entering your email address and clicking the Reset Password button. You will then receive an email that contains a secure link for resetting your password
If the address matches a valid account an email will be sent to __email__ with instructions for resetting your password
| Property | Value |
|---|---|
| Status | |
| Version | |
| Ad File | |
| Disable Ads Flag | |
| Environment | |
| Moat Init | |
| Moat Ready | |
| Contextual Ready | |
| Contextual URL | |
| Contextual Initial Segments | |
| Contextual Used Segments | |
| AdUnit | |
| SubAdUnit | |
| Custom Targeting | |
| Ad Events | |
| Invalid Ad Sizes |
Access provided by
The Cambrian explosion
Download started
- Download PDF Download PDF
Evidence of Cambrian diversity
The ediacaran period.

Creatures of the Cambrian

Fossilization during the Cambrian

Insights from fossils and genes

Where is the Cryogenian record?
Setting off the explosion.

Further Reading
Article metrics, related articles.
- Download Hi-res image
- Download .PPT
- Cancer Cell
- Cell Chemical Biology
- Cell Genomics
- Cell Host & Microbe
- Cell Metabolism
- Cell Reports
- Cell Reports Medicine
- Cell Stem Cell
- Cell Systems
- Current Biology
- Developmental Cell
- Molecular Cell
- American Journal of Human Genetics ( partner )
- Biophysical Journal ( partner )
- Biophysical Reports ( partner )
- Human Genetics and Genomics Advances ( partner )
- Molecular Plant ( partner )
- Molecular Therapy ( partner )
- Molecular Therapy Methods & Clinical Development ( partner )
- Molecular Therapy Nucleic Acids ( partner )
- Molecular Therapy Oncology ( partner )
- Plant Communications ( partner )
- Stem Cell Reports ( partner )
- Trends in Biochemical Sciences
- Trends in Cancer
- Trends in Cell Biology
- Trends in Ecology & Evolution
- Trends in Endocrinology & Metabolism
- Trends in Genetics
- Trends in Immunology
- Trends in Microbiology
- Trends in Molecular Medicine
- Trends in Neurosciences
- Trends in Parasitology
- Trends in Pharmacological Sciences
- Trends in Plant Science
- Cell Reports Physical Science
- Chem Catalysis
- Trends in Chemistry
- Cell Biomaterials
- Cell Reports Methods
- Cell Reports Sustainability
- STAR Protocols
- Nexus ( partner )
- The Innovation ( partner )
- Trends in Biotechnology
- Trends in Cognitive Sciences
- Submit article
- Multi-Journal Submission
- STAR Methods
- Sneak Peek – Preprints
- Information for reviewers
- Cell Symposia
- Consortia Hub
- Cell Press Podcast
- Cell Press Videos
- Coloring and Comics
- Cell Picture Show
- Research Arc
- About Cell Press
- Open access
- Sustainability hub
- Inclusion and diversity
- Help & Support
- Cell Press Careers
- Scientific job board
- Read-It-Now
- Recommend to Librarian
- Publication Alerts
- Best of Cell Press
- Cell Press Reviews
- Cell Press Selections
- Nucleus Collections
- SnapShot Archive
- For Advertisers
- For Recruiters
- For Librarians
- Privacy Policy
- Terms and Conditions
- Accessibility
The content on this site is intended for healthcare professionals and researchers across all fields of science.
We use cookies to help provide and enhance our service and tailor content. To update your cookie settings, please visit the Cookie settings for this site. All content on this site: Copyright © 2024 Elsevier Inc., its licensors, and contributors. All rights are reserved, including those for text and data mining, AI training, and similar technologies. For all open access content, the Creative Commons licensing terms apply.
- Privacy Policy
- Terms & Conditions
- Accessibility
- Help & Support

Session Timeout (2:00)
Your session will expire shortly. If you are still working, click the ‘Keep Me Logged In’ button below. If you do not respond within the next minute, you will be automatically logged out.
Thank you for visiting nature.com. You are using a browser version with limited support for CSS. To obtain the best experience, we recommend you use a more up to date browser (or turn off compatibility mode in Internet Explorer). In the meantime, to ensure continued support, we are displaying the site without styles and JavaScript.
- View all journals
- Explore content
- About the journal
- Publish with us
- Sign up for alerts
- Published: 18 April 2012
Formation of the ‘Great Unconformity’ as a trigger for the Cambrian explosion
- Shanan E. Peters 1 &
- Robert R. Gaines 2
Nature volume 484 , pages 363–366 ( 2012 ) Cite this article
10k Accesses
259 Citations
91 Altmetric
Metrics details
- Geodynamics
- Palaeontology
The transition between the Proterozoic and Phanerozoic eons, beginning 542 million years (Myr) ago, is distinguished by the diversification of multicellular animals and by their acquisition of mineralized skeletons during the Cambrian period 1 . Considerable progress has been made in documenting and more precisely correlating biotic patterns in the Neoproterozoic–Cambrian fossil record with geochemical and physical environmental perturbations 2 , 3 , 4 , 5 , but the mechanisms responsible for those perturbations remain uncertain 1 , 2 . Here we use new stratigraphic and geochemical data to show that early Palaeozoic marine sediments deposited approximately 540–480 Myr ago record both an expansion in the area of shallow epicontinental seas and anomalous patterns of chemical sedimentation that are indicative of increased oceanic alkalinity and enhanced chemical weathering of continental crust. These geochemical conditions were caused by a protracted period of widespread continental denudation during the Neoproterozoic followed by extensive physical reworking of soil, regolith and basement rock during the first continental-scale marine transgression of the Phanerozoic. The resultant globally occurring stratigraphic surface, which in most regions separates continental crystalline basement rock from much younger Cambrian shallow marine sedimentary deposits, is known as the Great Unconformity 6 . Although Darwin and others have interpreted this widespread hiatus in sedimentation on the continents as a failure of the geologic record, this palaeogeomorphic surface represents a unique physical environmental boundary condition that affected seawater chemistry during a time of profound expansion of shallow marine habitats. Thus, the formation of the Great Unconformity may have been an environmental trigger for the evolution of biomineralization and the ‘Cambrian explosion’ of ecologic and taxonomic diversity following the Neoproterozoic emergence of animals.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
185,98 € per year
only 3,65 € per issue
Buy this article
- Purchase on SpringerLink
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout
Similar content being viewed by others

Peak Cenozoic warmth enabled deep-sea sand deposition

A diverse Ediacara assemblage survived under low-oxygen conditions

Life before impact in the Chicxulub area: unique marine ichnological signatures preserved in crater suevite
Marshall, C. R. Explaining the Cambrian “Explosion” of animals. Annu. Rev. Earth Planet. Sci. 34 , 355–384 (2006)
Article ADS CAS Google Scholar
Maloof, A. C. et al. The earliest Cambrian record of animals and ocean geochemical change. Geol. Soc. Am. Bull. 122 , 1731–1774 (2010)
Canfield, D. E., Poulton, S. W. & Narbonne, G. M. Late-Neoproterozoic deep-ocean oxygenation and the rise of animal life. Science 315 , 92–95 (2007)
Dahl, T. W. et al. Devonian rise in atmospheric oxygen correlated to radiations of terrestrial plants and large predatory fish. Proc. Natl Acad. Sci. USA 107 , 17911–17915 (2010)
Gill, B. C. et al. Geochemical evidence for widespread euxinia in the later Cambrian ocean. Nature 469 , 80–83 (2011)
Yochelson, E. L. The Lipalian interval: a forgotten, novel concept in the geologic column. Earth Sci. Hist. 25 , 251–269 (2006)
Article Google Scholar
Avigad, D. et al. Mass-production of Cambro-Ordovician quartz-rich sandstone as a consequence of chemical weathering of Pan-African terranes: environmental implications. Earth Planet. Sci. Lett. 240 , 818–826 (2005)
Laird, M. G. in Geological Evolution of Antarctica (eds Thomson, M. R. A., Crame, J. A. & Thomson, J. W. ) 177–186 (Cambridge Univ. Press, 1991)
Brasier, M. D. The Lower Cambrian transgression and glauconite-phosphate facies in western Europe. J. Geol. Soc. Lond. 137 , 695–703 (1980)
Article CAS Google Scholar
Sears, J. W. & Price, R. A. Tightening the Siberian connection to western Laurentia. Geol. Soc. Am. Bull. 115 , 943–953 (2003)
Article ADS Google Scholar
Sloss, L. L. Sequences in the cratonic interior of North America. Geol. Soc. Am. Bull. 74 , 93–114 (1963)
Meyers, S. R. & Peters, S. E. A 56 million year rhythm in North American sedimentation during the Phanerozoic. Earth Planet. Sci. Lett. 303 , 174–180 (2011)
Cross, T. A. & Lessenger, M. A. in Innovative Applications of Petroleum Technology in the Rocky Mountain Region (eds Coalson, E. B., Osmond, J. C. & Williams, E. T. ) 183–203 (Rocky Mountain Association of Geologists, Denver, 1997)
Mortatti, J. & Probst, J. Silicate rock weathering and atmospheric/soil CO2 uptake in the Amazon basin estimated from river water geochemistry: seasonal and spatial variations. Chem. Geol. 197 , 177–196 (2003)
Millot, R. Gaillardet, J. Dupré, B. & Allegre, C. J. The global control of silicate weathering rates and the coupling with physical erosion: new insights from rivers of the Canadian Shield. Earth Planet. Sci. Lett. 196 , 83–98 (2002)
Robison, R. A. in Guidebook to the Geology of East Central Nevada (eds Boettcher, J. W. Jr & Sloan, W. W. ) 43–52 (Intermountain Association of Petroleum Geologists, Salt Lake City, 1960)
Ronov, A. B., Khain, V. E., Balukhovsky, A. N. & Seslavinsky, K. B. Quantitative analysis of Phanerozoic sedimentation. Sedim. Geol. 25 , 311–325 (1980)
Walker, L. J., Wilkinson, B. H. & Ivany, L. C. Continental drift and Phanerozoic carbonate accumulation in shallow-shelf and deep-marine settings. J. Geol. 110 , 75–87 (2002)
Ginsburg, R. N. Actualistic depositional models for the Great American Bank (Cambro-Ordovician). In Eleventh International Congress on Sedimentology , Abstracts of Papers 114 (International Association of Sedimentologists/McMaster University, 1982)
Google Scholar
Ridgwell, A. J., Kennedy, M. J. & Caldeira, K. Carbonate deposition, climate stability, and Neoproterozoic ice ages. Science 302 , 859–862 (2003)
Gaines, R. R. et al. Mechanism for Burgess Shale-type preservation. Proc. Natl Acad. Sci. USA http://dx.doi.org/10.1073/pnas.1111784109 (published online, 5 March 2012)
Coleman, M. L. Geochemistry of diagenetic non-silicate minerals: kinetic considerations. Phil. Trans. R. Soc. Lond. A 315 , 39–56 (1985)
Elrick, M. & Snider, A. S. Deep-water stratigraphic cyclicity and carbonate mud mound development in the Middle Cambrian Marjum Formation, House Range, Utah, U.S.A. Sedimentology 49 , 1021–1047 (2002)
Odin, G. S. & Matter, A. De glauconiarum origine. Sedimentology 28 , 611–641 (1981)
Chafetz, H. S. & Reid, A. Syndepositional shallow-water precipitation of glauconite minerals. Sedim. Geol. 136 , 29–42 (2000)
Keto, L. S. & Jacobson, S. B. Nd isotopic variations of Phanerozoic palaeoceans. Earth Planet. Sci. Lett. 90 , 395–410 (1988)
Lowenstein, T. K. et al. Oscillations in Phanerozoic seawater chemistry: evidence from fluid inclusions. Science 294 , 1086–1088 (2001)
Brennan, S. T., Lowenstein, T. K. & Horita, J. Seawater chemistry and the advent of biocalcification. Geology 32 , 473–476 (2004)
Petrychenko, O. Y., Peryt, T. M. & Chechei, E. I. Early Cambrian seawater chemistry from fluid inclusions in halite from Siberian evaporites. Chem. Geol. 219 , 149–161 (2005)
Erwin, D. H. et al. The Cambrian conundrum: early divergence and later ecological success in the early history of animals. Science 334 , 1091–1097 (2011)
Download references
Acknowledgements
We thank D. Canfield, P. Cohen, W. Fischer, S. Finnegan, N. Heim, A. Carroll and R. Dott for discussion, and N. Butterfield, M. Foote, E. Hammarlund, P. Myrow, B. Wilkinson, R. Wood, S. Holland for feedback on ideas. Fieldwork and analysis was aided by P. Burke, J. B. Caron, L. Curtin, F. Dwan, Z. Feng, P. Fenton, L. Finley-Blasi, X. Hou, J. Lackey, C. Qi, J. Peng, J. Tian, J. Vorhies, Y. Yang, X. Zhang and Y. Zhao. Work was supported by NSF EAR-0819931 (to S.E.P.) and EAR-1046233 and DUE-0942447 (to R.R.G.).
Author information
Authors and affiliations.
Department of Geoscience, University of Wisconsin, Madison, 53706, Wisconsin, USA
Shanan E. Peters
Geology Department, Pomona College, Claremont, 91711, California, USA
Robert R. Gaines
You can also search for this author in PubMed Google Scholar
Contributions
S.E.P. contributed Macrostrat-derived data, R.R.G. contributed sample-derived data. Both authors contributed to the development of ideas and writing.
Corresponding author
Correspondence to Shanan E. Peters .
Ethics declarations
Competing interests.
The authors declare no competing financial interests.
Additional information
Data for aspects of this analysis derive from Macrostrat ( http://macrostrat.org ).
Supplementary information
Supplementary information.
This file contains Supplementary Figures 1-8, Supplementary Tables 1-3 and additional references. (PDF 9479 kb)
PowerPoint slides
Powerpoint slide for fig. 1, powerpoint slide for fig. 2, powerpoint slide for fig. 3, powerpoint slide for fig. 4, rights and permissions.
Reprints and permissions
About this article
Cite this article.
Peters, S., Gaines, R. Formation of the ‘Great Unconformity’ as a trigger for the Cambrian explosion. Nature 484 , 363–366 (2012). https://doi.org/10.1038/nature10969
Download citation
Received : 29 June 2011
Accepted : 16 February 2012
Published : 18 April 2012
Issue Date : 19 April 2012
DOI : https://doi.org/10.1038/nature10969
Share this article
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
This article is cited by
Glauconite facies developed sequentially in the abu tartur plateau (egypt) during the late cretaceous.
- Abdalla M. El Ayyat
- Samia El-Helaly
- Mostafa R. Abukhadra
Euro-Mediterranean Journal for Environmental Integration (2024)
Secular craton evolution due to cyclic deformation of underlying dense mantle lithosphere
- Xiaotao Yang
Nature Geoscience (2023)
Reappraisal of the Neoproterozoic to middle Paleozoic fossils of North Korea and its tectonic implication
- Jikhan Jung
- Tae-Yoon S. Park
Geosciences Journal (2023)
Diversified calcimicrobes in dendrolites of the Zhangxia Formation, Miaolingian Series (Middle Cambrian) of the North China craton
- Ming-Xiang Mei
- Muhammad Riaz
Journal of Palaeogeography (2021)
Current understanding on the Cambrian Explosion: questions and answers
- Xingliang Zhang
PalZ (2021)
By submitting a comment you agree to abide by our Terms and Community Guidelines . If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
Quick links
- Explore articles by subject
- Guide to authors
- Editorial policies
Sign up for the Nature Briefing newsletter — what matters in science, free to your inbox daily.

- Access through your organization
- Purchase PDF
Article preview
Introduction, section snippets, references (165), cited by (100).

Gondwana Research
Gr focus review triggers for the cambrian explosion: hypotheses and problems.
- • The developmental system of bilaterians was established before their divergence.
- • This, in turn, suggests that the Cambrian explosion require environmental triggers.
- • The Cambrian explosion is the initial formation of metazoan-dominated ecosystem.
Graphical abstract

- Download: Download full-size image
Hypothetical triggers
Conclusions, outlook for future research, acknowledgments, what sponges can tell us about the evolution of developmental processes, the mountains that triggered the late neoproterozoic increase in oxygen: the second great oxidation event, geochimica et cosmochimica acta, true polar wander, a supercontinental legacy, earth and planetary science letters, true polar wander and supercontinents, tectonophysics, the cambrian evolutionary ‘explosion’: decoupling cladogenesis from morphological disparity, biological journal of the linnean society, evaporites and the salinity of the ocean during the phanerozoic: implications for climate, ocean circulation and life, palaeogeography, palaeoclimatology, palaeoecology, a methane fuse for the cambrian explosion: carbon cycles and true polar wander, comptes rendus geosciences, temperature and salinity history of the precambrian ocean: implications for the course of microbial evolution, evolution of the composition of seawater through geologic time, and its influence on evolution of life, an outline of the palaeongeographic evolution of the australasian region since the beginning of the neoproterozoic, earth-science reviews, models on snowball earth and cambrian explosion: a synopsis, giant deep-sea protist produces bilaterian-like traces, current biology, the vendian (ediacaran) in the geological record: enigmas in geology's prelude to the cambrian explosion, the new animal phylogeny: reliability and implications, proceedings of the national academy of sciences of the united states of america, facilitation cascade drives positive relationship between native biodiversity and invasion success, extinction of cloudina and namacalathus at the precambrian–cambrian boundary in oman, proterozoic ocean chemistry and evolution: a bioinorganic bridge, bayesian models of episodic evolution support a late precambrian explosive diversification of the metazoa, molecular biology and evolution, the dawn of bilaterian animals: the case of acoelomorph flatworms, back in a time: a new systematic proposal for the bilateria, philosophical transactions of the royal society of london b, the genetic response to snowball earth: role of hsp90 in the cambrian explosion, the segmented urbilateria: a testable scenario, integrative and comparative biology, origins and early evolution of predation, paleontological society papers, predatorial borings in late precambrian mineralized exoskeletons, evolution of microrna diversity and regulation in animals, nature reviews genetics, a model for calcium, magnesium and sulfate in seawater over phanerozoic time, american journal of science, animals (metazoa), the cambrian substrate revolution, neoproterozoic ‘snowball earth’ glaciations and the evolution of altruism, did supercontinental amalgamation trigger the “cambrian explosion”, seawater chemistry and the advent of biocalcification, testing the cambrian explosion hypothesis by using a molecular dating technique, the earliest fossil record of the animals and its significance, philosophical transactions of the royal society b, plankton ecology and the proterozoic–phanerozoic transition, paleobiology, cambrian food webs, oxygen, animals and oceanic ventilation: an alternative view, terminal developments in ediacaran embryology, formation of supercontinents linked to increases in atmospheric oxygen, nature geoscience, the early history of atmospheric oxygen: homage to robert m. garrels, annual review of earth and planetary sciences, animal evolution, bioturbation, and the sulfate concentration of the oceans, late proterozoic rise in atmospheric oxygen concentration inferred from phylogenetic and sulphur-isotope studies, late-neoproterozoic deep-ocean oxygenation and the rise of animal life, ferruginous conditions dominated later neoproterozoic deep-water chemistry, endless forms most beautiful: the new science of evo devo and the making of the animal kingdom, from dna to diversity: molecular genetics and the evolution of animal design, why o 2 is required by complex life on habitable planets and the concept of planetary “oxygenation time”, astrobiology, the advent of hard-part structural support among the ediacara biota: ediacaran harbinger of a cambrian mode of body construction, beginnings of biospheric evolution and their biogeochemical consequences, the cambrian “explosion”: slow-fuse or megatonnage, darwin's dilemma: the realities of the cambrian ‘explosion’, philosophy transactions of the royal society london b, marine oxygenation, lithistid sponges, and the early history of paleozoic skeletal reefs, the lower ordovician fezouata konservat-lagerstätte from morocco: age, environment and evolutionary perspectives, birth and early evolution of metazoans, the temporal and environmental context of early animal evolution: considering all the ingredients of an “explosion”, atmospheric evolution on inhabited and lifeless worlds, the latest ediacaran wormworld fauna: setting the ecological stage for the cambrian explosion.

- Faculty Experts
- Events Calendar
Evidence for a geologic trigger of the Cambrian explosion
The oceans teemed with life 600 million years ago, but the simple, soft-bodied creatures would have been hardly recognizable as the ancestors of nearly all animals on Earth today.
Then something happened. Over several tens of millions of years — a relative blink of an eye in geologic terms — a burst of evolution led to a flurry of diversification and increasing complexity, including the expansion of multicellular organisms and the appearance of the first shells and skeletons.
The Great Unconformity is visible in the Grand Canyon at the base of a rock cliff above where the canyon walls slope down to the Colorado River. The flat-lying layered sedimentary rocks of the 525-million-year-old Cambrian Tapeats Sandstone rest on metamorphic rocks of the 1,740-million-year-old Vishnu Schist. (Photo: Jack Share)
The results of this Cambrian explosion are well documented in the fossil record, but its cause — why and when it happened, and perhaps why nothing similar has happened since — has been a mystery.
New research shows that the answer may lie in a second geological curiosity — a dramatic boundary, known as the Great Unconformity, between ancient igneous and metamorphic rocks and younger sediments.
“The Great Unconformity is a very prominent geomorphic surface and there’s nothing else like it in the entire rock record,” says Shanan Peters , a geoscience professor at the University of Wisconsin–Madison who led the new work. Occurring worldwide, the Great Unconformity juxtaposes old rocks, formed billions of years ago deep within the Earth’s crust, with relatively young Cambrian sedimentary rock formed from deposits left by shallow ancient seas that covered the continents just a half billion years ago.
Named in 1869 by explorer and geologist John Wesley Powell during the first documented trip through the Grand Canyon, the Great Unconformity has posed a longstanding puzzle and has been viewed — by Charles Darwin, among others — as a huge gap in the rock record and in our understanding of the Earth’s history.
But Peters says the gap itself — the missing time in the geologic record — may hold the key to understanding what happened.
In the April 19 issue of the journal Nature, he and colleague Robert Gaines of Pomona College report that the same geological forces that formed the Great Unconformity may have also provided the impetus for the burst of biodiversity during the early Cambrian.
A Cambrian trilobite, with a shell made of calcium carbonate. (Photo: Shanan Peters)
“The magnitude of the unconformity is without rival in the rock record,” Gaines says. “When we pieced that together, we realized that its formation must have had profound implications for ocean chemistry at the time when complex life was just proliferating.”
“We’re proposing a triggering mechanism for the Cambrian explosion,” says Peters. “Our hypothesis is that biomineralization evolved as a biogeochemical response to an increased influx of continental weathering products during the last stages in the formation of the Great Unconformity.”
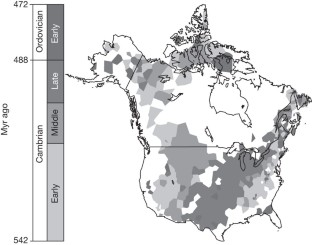
Peters and Gaines looked at data from more than 20,000 rock samples from across North America and found multiple clues, such as unusual mineral deposits with distinct geochemistry, that point to a link between the physical, chemical, and biological effects.
During the early Cambrian, shallow seas repeatedly advanced and retreated across the North American continent, gradually eroding away surface rock to uncover fresh basement rock from within the crust. Exposed to the surface environment for the first time, those crustal rocks reacted with air and water in a chemical weathering process that released ions such as calcium, iron, potassium, and silica into the oceans, changing the seawater chemistry.
The basement rocks were later covered with sedimentary deposits from those Cambrian seas, creating the boundary now recognized as the Great Unconformity.
Evidence of changes in the seawater chemistry is captured in the rock record by high rates of carbonate mineral formation early in the Cambrian, as well as the occurrence of extensive beds of glauconite, a potassium-, silica-, and iron-rich mineral that is much rarer today.
The influx of ions to the oceans also likely posed a challenge to the organisms living there. “Your body has to keep a balance of these ions in order to function properly,” Peters explains. “If you have too much of one you have to get rid of it, and one way to get rid of it is to make a mineral.”
The fossil record shows that the three major biominerals — calcium phosphate, now found in bones and teeth; calcium carbonate, in invertebrate shells; and silicon dioxide, in radiolarians — appeared more or less simultaneously around this time and in a diverse array of distantly related organisms.
The time lag between the first appearance of animals and their subsequent acquisition of biominerals in the Cambrian is notable, Peters says. “It’s likely biomineralization didn’t evolve for something, it evolved in response to something — in this case, changing seawater chemistry during the formation of the Great Unconformity. Then once that happened, evolution took it in another direction.” Today those biominerals play essential roles as varied as protection (shells and spines), stability (bones), and predation (teeth and claws).
Together, the results suggest that the formation of the Great Unconformity may have triggered the Cambrian explosion.
“This feature explains a lot of lingering questions in different arenas, including the odd occurrences of many types of sedimentary rocks and a very remarkable style of fossil preservation. And we can’t help but think this was very influential for early developing life at the time,” Gaines says.
Far from being a lack of information, as Darwin thought, the gaps in the rock record may actually record the mechanism as to why the Cambrian explosion occurred in the first place, Peters says.
“The French composer Claude Debussy said, ‘Music is the space between the notes.’ I think that is the case here,” he says. “The gaps can have more information, in some ways, about the processes driving Earth system change, than the rocks do. It’s both together that give the whole picture.”
The work was supported by the National Science Foundation.
Tags: College of Letters & Science , research
- Search Menu
- Sign in through your institution
- Advance articles
- Author Guidelines
- Open Access
- About Integrative and Comparative Biology
- About the Society for Integrative and Comparative Biology
- Editorial Board
- Advertising and Corporate Services
- Journals Career Network
- Self-Archiving Policy
- Dispatch Dates
- Journals on Oxford Academic
- Books on Oxford Academic
Article Contents
Evolution of animal body plans, molecular clocks, molecular dates and explosive radiations, explosive radiations and the molecular clock, molecular dating with variable rates, metazoan molecular dates: where to next.
- < Previous
What can DNA Tell us About the Cambrian Explosion? 1
- Article contents
- Figures & tables
- Supplementary Data
Lindell Bromham, What can DNA Tell us About the Cambrian Explosion?, Integrative and Comparative Biology , Volume 43, Issue 1, February 2003, Pages 148–156, https://doi.org/10.1093/icb/43.1.148
- Permissions Icon Permissions
Molecular data is ideal for exploring deep evolutionary history because of its universality, stochasticity and abundance. These features provide a means of exploring the evolutionary history of all organisms (including those that do not tend to leave fossils), independently of morphological evolution, and within a statistical framework that allows testing of evolutionary hypotheses. In particular, molecular data have an important role to play in examining hypotheses concerning the tempo and mode of evolution of animal body plans. Examples are given where molecular phylogenies have led to a re-examination of some fundamental assumptions in metazoan evolution, such as the immutability of early developmental characters, and the evolvability of bauplan characters. Molecular data is also providing a new and controversial timescale for the evolution of animal phyla, pushing the major divisions of the animal kingdom deep into the Precambrian. There have been many reasons to question the accuracy and precision of molecular date estimates, such as the failure to account for lineage-specific rate variation and unreliable estimation of rates of molecular evolution. While these criticisms have been largely countered by recent studies, one problem has remained a challenge: could temporal variation in the rate of molecular evolution, perhaps associated with “explosive” adaptive radiations, cause overestimation of diversification dates? Empirical evidence for an effect of speciation rate, morphological evolution or ecological diversification on rates of molecular evolution is examined, and the potential for rate-variable methods for molecular dating are discussed.
The questions asked by George Gaylord Simpson in his 1944 , “The Tempo and Mode of Evolution,” concerning the size of mutations, the pace of morphological change and the apparent discontinuous origins of taxa in the fossil record, are far from resolved. Indeed, they are being debated more strongly than ever, because of the growing conviction amongst many biologists that observations from developmental biology and palaeontology are inconsistent with the Neodarwinian hypothesis championed by Simpson. The origin of the animal phyla has been a key case study in the tempo and mode of evolution.
The near-simultaneous appearance in the early Cambrian of the first recognizable members of many animal phyla, at high diversity and with few clear precursors, has been attributed by many researchers an incomplete fossil record ( e.g., Darwin, 1859 ). However, the description of late Precambrian body fossils and traces from around the world, from which members of diverse Cambrian phyla such as arthropods are conspicuously absent, has made arguments for a missing history less convincing. More broadly, analyses of fossil ranges across a wide range of taxa have led to increasing faith in the fossil record as an accurate record of tempo and mode of evolution ( e.g., Benton et al. , 2000 ). Such analyses have led to the suggestion that rather than a steady accumulation of biological diversity, the global biota has been fundamentally shaped by a series of major events, mass extinctions of taxa followed by explosive radiations of new taxa. The first explosive radiation—the Cambrian explosion—is particularly important, as it has been argued that the diversity of animal body plans was achieved soon after their origin, and that virtually no new body-plans have evolved since.
The apparently sudden origin of animal phyla has contributed to the view that phyla represent a fundamental level of organization. In particular, phyla are held to be defined primarily by a set of discrete body plan characters, rather than simply differing at a large number of continuously varying traits. Just as species are considered by many to represent a fundamental evolutionary unit, not just an arbitrary division of biological diversity, phyla are considered by many not simply as a taxonomic division, but as a reflection of an underlying biological structure.
The origin of animal phyla (both their sudden appearance in the fossil record and the discontinuous variation in bodyplan traits between phyla) has also been the focus of research in the relatively young field of evolutionary development (Evo-Devo). The characterization of homeotic mutants, that alter the development of whole structures such as limbs or eyes, has led some researchers to suggest that the differences in body plan between animal phyla could have arisen through relatively few genetic changes. In particular, the Hox genes (and related genes) have been implicated as controllers of body plan characters, and the differences between phyla have been been attributed to variation in the number and expression of Hox-like genes. Furthermore, the Cambrian explosion has been interpreted as the point at which Hox clusters formed, and were subsequently canalized so that no new body plans formed after that time ( e.g., Valentine et al. , 1999 ).
So these three observations—the sudden appearance of animal phyla in the fossil record, the discontinuity between animal body plans, and mutations that bring about discrete body-plan-like changes—have been combined in the hypothesis that the evolution of animal body plans did not occur by the gradual accumulation of small genetic differences, but by relatively few developmental changes with large phenotypic effects. Because this hypothesis offers a direct challenge to the Neodarwinian view of evolution (evolutionary change by the gradual accumulation of adaptive changes), the importance of these interpretations of the Cambrian explosion go beyond explaining a single evolutionary radiation.
The Neodarwinian hypothesis does not assume constant rates of evolutionary change, but it does assume uniformity of process—that all evolutionary change arises by the same basic mechanism. Darwin's (1859) hypothesis provided a unifying theory for biological diversity because his mechanism for evolution was explicity uniformitarian. He followed the revolutionary works of Charles Lyell (1830) in which the dramatic geological changes of the past—such as mountain building or sea level changes—were explained in terms of the cumulative effect of processes witnessed in operation today, such as sedimentation or uplift. Darwin explained the extraordinary changes in species over time using a simple mechanism operating at the level of populations: the same process that causes relatively modest differences over observable periods produces major changes over much longer timescales. In other words, macroevolution (formation of higher taxa) is simply the cumulative effect of microevolution (population genetic changes). The new view of evolution challenges this assumption, by proposing that the evolution of animal body-plans occurred by a different process than that which modifies existing body parts. If the differences between animal phyla must be explained by a process that operated in the early Cambrian, but not before or since, then this is a serious challenge to the uniformitarian principles upon which evolutionary biology has been based for the past century and a half. It is therefore very important that the hypothesis is thoroughly tested.
Molecular data can contribute to this debate by providing a means of inferring phylogeny independently of morphological characters, and a timescale independent of the fossil record. The key features of molecular data that make it so useful for assessing tempo and mode of evolution are universality, abundance and stochasticity. DNA data is directly comparable across all extant organisms, including those that do not fossilize well such as soft bodied taxa, and contains information for the entire history of every lineage. DNA sequence data provides an abundant data source, with thousands of independently evolving characters in even the smallest genome. DNA sequences evolve in a predominantly stochastic manner because, although every heritable change to the phenotype must be associated with a change in the genome, the reverse is not true: most changes to the genome are apparently unconnected to phenotypic change. Many changes occur in non-translated DNA sequences, or at sites that do not make a functional change to a gene product. Because most DNA substitutions will have little effect on fitness, the rate of accumulation of genetic change should increase as a function of time. These features of molecular data provide the basis for independent tests of hypotheses concerning the tempo and mode of metazoan evolution generated from morphological phylogeny and fossil evidence.
The hypothesis that body plans were formed in the Cambrian Explosion then canalized predicts that there are critical characters that not only define phyla, but were instrumental in their formation ( e.g., Carroll, 1995 ; Erwin, 1993 ; Valentine et al. , 1999 ). This contrasts to the Neodarwinian perspective that the apparent origin of phyla in the Cambrian is simply a reflection of the amount of time needed to accumulate sufficient differences to be considered separate phyla ( e.g., Williams, 1992 ). The claim that critical body plan characters formed in the Cambrian, but not since, can be tested directly from fossil evidence, which provides evidence of observable bodyplan characters present before, during and after the Cambrian explosion ( e.g., Budd and Jensen, 2000 ), or indirectly by inference of the relative timing of character evolution from phylogenies. However, assessing the relative timing of evolution of body plan characters from phylogenies constructed from morphological data risks a circular argument. If one of the assumptions on which the phylogeny is based is that a set of characters are so fundamental that they will evolve only once, and therefore define major groups, then the resulting phylogeny cannot be used to test the same assumption. The stochastic nature of molecular change makes it ideal for providing an independent source of phylogenetic information to test these claims about the tempo and mode of morphological evolution.
The deuterostomes provide an illustrative example of the use of molecular phylogenies to examine the evolution of body plan characters. The superphylum Deuterostomia contains the diverse phyla Chordata and Echinodermata and the minor phyla Hemichordata and Urochordata. Although united by early developmental characteristics, echinoderms and chordates have strikingly different adult body plans. The head-and-tail chordate body plan is characterized by a pharynx, dorsal nerve cord and post-anal tail. The echinoderm body plan has pentameral symmetry, a water vascular system and a ring-shaped nervous system. Hemichordates have traditionally been allied with the chordates ( Fig. 1a ) on the basis of shared adult body plan features, particularly the pharynx and dorsal nerve chord. Molecular phylogenies ( Fig. 1b ) have suggested that hemichordates are in fact a sister lineage to the echinoderms ( e.g., Castresana et al. , 1998 ; Holland et al. , 1991 ). The morphological phylogeny gave few clues of the body plan of the ancestor to all deuterostome phylogeny ( Fig. 1a ). But if hemichordates are the sister group to echinoderms, then the deuterostome ancestor must have had chordate body plan characters including a pharynx and dorsal nerve chord, because these characters are present in both major branches of the deuterosomes ( Bromham and Degnan, 1999 ). So molecular data suggest that the echinoderm lineage, with its distinctive pentameral body plan, evolved from an ancestor that had the key features of the head-and-tail chordate body plan. It also implies that aspects of the chordate body plan did not evolve as a single complex. Instead, the postanal muscularized tail of chordates evolved after the pharynx ( Bromham and Degnan, 1999 ; Hinman and Degnan, 2000 ). Whatever timescale this change in body plan characters occurred on, it suggests that body plans of metazoan phyla were not formed once then canalized, but were able to evolve in stepwise fashion from one body plan to another. Animal body plans are therefore not immutable.
Early developmental characters have also been considered to be relatively immutable. The assumption that the earlier a character is in the developmental hierarchy, the less it may be modified in evolution, has provided the basis for constructing phylogenies of the animal kingdom ( e.g., Haeckel, 1879 ). By providing phylogenies independent of developmental characters, DNA sequence data have allowed these assumptions to be re-examined. In some cases, this has supported the phylogenetic significance of developmental characters, such as the superphylum Spiralia which is characterized by a pattern of spiral cleavage in the early embryo. In other cases, molecular phylogenies suggest that some developmental characters are less constrained than is often assumed. The minor phyla of Spiralia may provide a illustrative case. The phylum Sipuncula (acorn worms) has been considered allied to the molluscs on the basis of a pattern of cells in early spiral cleavage of the embryo, known as the molluscan cross. But some molecular phylogenies ( e.g., Peterson and Eernisse, 2001 ; Boore and Staton, 2002 ) place Sipuncula with Annelida, a grouping that implies that molluscan cross pattern of early spiral cleavage evolved several times independently. The phylogenetic position of the Sipuncula is still unclear, but whether or not the alliance between Sipuncula and annelids is confirmed by future phylogenetic analyses (both molecular and morphological), this initial result should lead to careful scrutiny of character coding. The perception that early developmental characters are fundamental may have led to bias in coding some developmental patterns, which may form a continuous variation in early cell sizes and patterns, rather than a number of discrete, distinctive developmental patterns (see Jenner, 2003 ).
Molecular phylogenies of Metazoa suggest that even fundamental developmental and body plan characters can evolve along the metazoan phylogeny, rather than being fixed at the base of the radiation. Clearly the phylogenetic hypotheses generated from molecular data need to be thoroughly tested, particularly as at present most metazoan phylogenies are based on a single gene, 18s rRNA. It may be that some of these phylogenetic hypotheses are incorrect. But by providing a source of phylogenetic information independent of morphological and developmental characters, molecular phylogenies offer a way of testing traditional assumptions about the evolutionary lability of traits, and will in many cases lead to a reexamination of the characters on which systematic divisions of Metazoa have been based.
The stochastic nature of DNA sequence evolution leads to the prediction that substitutions should increase as a function of time. This prediction is broadly borne out: in general, the more distantly related two species are, the more sites in a DNA or protein sequence differ between them. In many cases, if genetic distance is plotted against time, a linear relationship is revealed ( e.g., Fleischer et al. , 1998 ; Runnegar, 1982 ; Zuckerkandl and Pauling, 1965 ), suggesting that genetic data can be used to predict divergence times. But molecular clock estimates for the origin of metazoan lineages are at odds with the timing of appearance of metazoan phyla in the fossil record. While molecular clock estimates vary greatly, all estimates for the major divisions of the Metazoa ( e.g., between bilaterians and deuterostomes) are all at least 55 Ma before the first unambiguous metazoan body fossils in the early Cambrian, and more than 25 Ma before the first multicellular animals in the Vendian (see Bromham and Hendy, 2000 ). Most molecular dates are one hundred million years or more before the Cambrian explosion.
There are a number of possible explanations of the discrepancy between molecular and morphological dates of the metazoan radiation. Firstly, the gap between the molecular dates and the first fossils may be due to incompleteness of the fossil record, such that the early evolutionary history of the metazoans is obscured. This is difficult to reconcile with the apparent increase in complexity of body and trace fossils across the latest Proterozoic and earliest Phanerozoic. Secondly, molecular and paleaontological dates may be effectively measuring different things, if lineage origination is disconnected from the evolution of morphological divergence. It has been suggested that the basal splits of the metazoan tree occurred in the late Proterozoic, but these lineages persisted in relatively low diversity until the early Cambrian. In this case, it may be that the predominance of molecular date estimates for basal splits (such as the protostome–deuterostome split) is responsible for the huge disparity between molecular and fossil dates, and that dates for “shallower” splits will be closer to the Cambrian. This view raises important questions about the tempo and mode of evolution, as it requires that the early metazoan lineages persisted in some kind of evolutionary stasis even though they had all of the elements of the body plan “toolkit” that have been considered by many to have been key innovations of the metazoan radiation. Thirdly, the discrepancy between the molecular and palaeontological dates might be due to a systematic bias in molecular clock estimates that results in consistent overestimation of the date of divergence of animal phyla.
There are a number of reasons to be worried about the accuracy and precision of molecular dates for the metazoan radiation:
Failure to account for rate variation: Most molecular dating techniques assume that the rate of molecular evolution is approximately the same in all taxa ( e.g., Lynch, 1999 ; Nei et al. , 2001 ). Since lineages can vary in rate of molecular evolution, various “clock tests” have been used to identify genes for which rates do not vary between lineages ( Wang et al. , 1999 ), such as the relative rates test ( Wu and Li, 1985 ) or the Tajima test ( Tajima, 1993 ). These tests have low power for the type of sequences typically used in molecular clock studies ( Bromham et al. , 2000 ). Using saturated sequences or distant outgroups (such fungi as an outgroup to Metazoa) also reduces the power of relative rates tests to detect rate variation ( e.g., Robinson et al. , 1998 ). Even relatively high levels of rate variation between lineages may not be detected by clock tests, leading to an erroneous impression of rate constancy. Undetected rate variation can lead to consistently overestimated dates of divergence ( Bromham et al. , 2000 ).
Poor estimation of branch length: selection of substitution model is a critical part of molecular dating. For example, failure to account for variation in substitution rate between sites in the sequence will result in inaccurate estimation of branch length. In the case of internal calibrations (where the calibration date is younger than the node for which the molecular date is estimated), this will generally shorten branches and thus make dates too early ( Ayala et al. , 1998 ): the opposite is likely to be true for external calibrations (where the calibration date is older than the estimated node). Similarly, exclusion of invariant sites ( Lynch, 1999 ) is likely to bias estimates of both substitution rates and branch lengths.
Misleading confidence intervals: Molecular date estimates are sometimes presented with confidence intervals that represent the standard deviation of a number of estimates (commonly from different genes). This is a reflection of difference between estimates, not of the accuracy of the estimates per se. For example, erroneous estimates that all share the same measurement bias might be represented by a mean with narrow confidence intervals. In this way, collating large numbers of genes can give a false sense of confidence in molecular date estimates. Giving equal weight to all genes also falsely reduces confidence intervals, because it does not account for the effect of sequence length, substitution rate, and degree of rate variation across sites on the precision of date estimation. In addition, care must be taken to consider whether the estimates are truly independent before mean estimates are calculated ( Bromham et al. , 1998 ). Ideally, confidence intervals should reflect not only the difference between estimates obtained from different genes or different calibration dates, but should also reflect lineage-specific rate variation and the “sloppiness” of the molecular clock (the random distribution of the time interval between substitutions), both of which add a great deal of imprecision to molecular date estimates (see Rambaut and Bromham, 1998 ).
Poor calibration dates: While some studies have aimed to use as many fossil calibration dates as possible, an alternative strategy has been use a single calibration date deemed to be well supported. The accuracy of one commonly used calibration date—310 Myr for the split between reptiles and mammals ( Gu, 1998 ; Wang et al. , 1999 )—has been questioned ( Lee, 1998 ). More broadly, the strategy of using rates calculated for a specific, potentially unrepresentative, group—particularly vertebrates—to date the rest of the metazoan tree ( Feng et al. , 1997 ; Wang et al. , 1999 ) has been criticized ( e.g., Lynch, 1999 ). Even “local” molecular clocks should be rigorously tested, as closely related lineages can vary substantially in substitution rate ( e.g., Bromham, 2002 ; Bromham et al. , 1996 ; Mooers and Harvey, 1994 ). Calibrating rates on other molecular clock estimates ( e.g., using estimate of primate-rodent at 100 Myr to date metazoan divergences: Gu, 1998 ; Wang et al. , 1999 ) is an ill-advised strategy, potentially compounding the error of earlier molecular clock estimates.
Regression through non-independent points: In some studies, calibration rates are calculated by taking the molecular distance between a number of pairs of taxa, each with a different calibration point, and extrapolating a linear relationship between genetic distance and time ( e.g., Doolittle et al. , 1996 ; Wray et al. , 1996 ). While these regressions of distance against time have the advantage of providing an instinctive test of the molecular clock, they are usually flawed due to the inclusion of non-independent points ( Lynch, 1999 ). The inclusion of the same branch-lengths in many datapoints ( Fig. 2 ) is likely to inflate the apparent association between branch-length and time, therefore giving false confidence in the “clock-like” evolution of a sequence.
Reliance on few sequences: 18s rRNA has dominated attempts to reconstruct molecular phylogenies for metazoans, and so has been used in many molecular dating studies ( e.g., Bromham et al. , 1998 ; Wray et al. , 1996 ). If this sequence gives misleading results, then it could influence the many studies that rely on 18s rRNA sequences ( Abouheif et al. , 1998 ). Phylogenetic inference from rRNA genes is complicated by the secondary structure of the RNA product: for example, no substitution model used in molecular phylogenetics accounts for paired substitutions in stem sites, and so the number of independent molecular changes may be overestimated for these sites, potentially producing molecular dates that are too old ( Bromham et al. , 1998 ).
However, none of these criticisms is sufficient to explain the Precambrian molecular dates for major metazoan divergences, because studies that overcame these problems have produced similar results. The accuracy of branch length estimation has been improved by using long sequences and methods that allow for variation in substitution rates across sites and between lineages ( Rambaut and Bromham, 1998 ). Biases due to specific genes or calibration dates have been countered by studies that use a range mitochondrial and nuclear protein-coding genes and many non-vertebrate calibrations ( Ayala et al. , 1998 ; Bromham et al. , 1998 ; Feng et al. , 1997 ; Gu, 1998 ; Wang et al. , 1999 ). And yet all of these analyses point to Precambrian divergences of major metazoan lineages.
One problem with molecular clocks that is more difficult to address is the potential for concerted changes in rate of molecular evolution over time. If lineages can have consistently different average substitution rates, this implies that rates evolve along phylogenies. This could create temporal patterns of substitution rates that would be difficult to detect, and hard to model. This is particularly worrying given that substitution rates can be correlated with species traits, such as life history or population dynamics ( e.g., Bromham, 2002 ; Bromham et al. , 1996 ; Johnson and Seger, 2001 ; Martin and Palumbi, 1993 ; Mooers and Harvey, 1994 ). A concerted evolutionary change in such species traits could generate temporal patterns in rates across many lineages, which could produce consistently misleading molecular date estimates ( Bromham, 2003 ). This problem can be illustrated with an example taken from another “explosive” radiation which mirrors the controversy over molecular dates for the metazoan radiation.
Modern mammal orders appear suddenly in the early Tertiary fossil record, but molecular date estimates put the the major mammalian divergences deep into the Cretaceous, long before the final extinction of the dinosaurs ( Kumar and Hedges, 1998 ; Madsen et al. , 2001 ). So molecular dates have been used to challenge the notion that the extinction of the dinosaurs was the key determinant of the radiation of modern mammals and birds, as they filled the niches left vacant by dinosaurs. This same pattern of early Tertiary fossils but Cretaceous molecular dates is repeated for the evolution of modern bird orders ( Cooper and Fortey, 1998 ; Cooper and Penny, 1997 ). Is this repeated pattern of molecular dates twice as old as an explosion of diversity in the fossil record due to systematic biases in the fossil record, or is there some aspect of explosive radiations that could lead to consistent overestimation of molecular dates?
Rates of molecular evolution can vary substantially between mammalian lineages ( e.g., Gillespie, 1991 ; Li et al. , 1996 ; Yang and Nielsen, 1998 ). This variation will not always be detected by “clock tests” such as the relative rates test, likelihood ratio tests, or the Tajima test ( Bromham et al. , 2000 ; Robinson et al. , 1998 ; Scherer, 1989 ). Undetected lineage-specific rate variation can result in consistent overestimation of molecular date estimates ( Bromham et al. , 2000 ). Furthermore, variation in rate of molecular evolution appears to be associated with body size in mammals ( Bromham et al. , 1996 ). This could be important for the accuracy of molecular dates of the mammalian radiation because virtually all modern mammal orders increased in average body size from their first appearance in the early Tertiary to the more diverse members of the order in the Eocene. If smaller species have faster rates of molecular evolution, then an increase in average body size during the mammalian radiation could have led to a concerted slowdown in rates of molecular evolution across many lineages. This slowdown would be difficult to detect, but could result in consistent overestimation of molecular dates for divergences between mammalian orders ( Bromham, 2003 ).
It is important to note that this line of reasoning does not prove that mammalian molecular dates are wrong. The molecular dates for mammalian divergences may be broadly correct, and the discrepancy with fossil data could be due to geographical biases in the late Cretaceous terrestrial record, a hypothesis supported by molecular phylogenetic analyses that place the origins of modern mammals in Gondwana ( e.g., Madsen et al. , 2001 ; Penny et al. , 1999 ). It is intended simply as an illustration of the possibility of consistent bias in molecular date estimates arising from patterns of rate of molecular evolution across lineages. Could a similar pattern account for the discrepancy between paleaontological and molecular dates for other explosive radiations? The body size pattern is unlikely to provide a general explanation—for example, the radiation of modern birds was not obviously marked by an increase in average body size. The pattern of body size evolution at the base of the metazoan radiation is unknown, and there have been no systematic studies of correlates of rate variation in invertebrates, so it is not known if life history influences molecular evolution in the majority of metazoan lineages.
Are there other determinants of rate of molecular evolution that could cause systematic errors in the molecular dates for the metazoan radiation? Adaptive radiations are characterized by several evolutionary processes that might influence rates of molecular evolution: increased speciation rate, rapid morphological change, and directional change in species characteristics.
Molecular evolution has been considered to be largely independent of the pace of phenotypic evolution, based on theoretical ( Kimura, 1983 ) and empirical ( Papadopoulos et al. , 1999 ) studies. Most substitutions are expected to be effectively neutral ( Kimura, 1983 ; Ohta, 1993 ), their fate determined by chance rather than selection, and the number of genes affected by any given selection event will be vanishingly small compared to the total genome size. However, a recent study suggesting an association between molecular and morphological branch lengths ( Omland, 1997 ) has been used to suggest that rapid phenotypic evolution associated with explosive radiations should be accompanied by faster molecular rates ( Archibald, 1999 ; Conway Morris, 1998 ; Lee, 1998 ). If true, such a relationship would have serious implications for using molecular clocks to date adaptive radiations. However, this study ( Omland, 1997 ) was limited by few datasets, and statistical flaws in the experimental design that could have inflated the probability of an artefactual association between morphological and molecular branch lengths for a number of phylogenies. A study that used three new methods designed to overcome these biases on thirteen “total evidence” vertebrate datasets found no evidence of association between morphological and molecular rates of change ( Bromham et al. , 2002 ). Clearly this relationship must be examined further before claims for rapid morphological evolution speeding the molecular clock in explosive radiations can be supported. Similarly, although an association between net speciation rate and substitution rate has been observed for three genes for angiosperms ( Barraclough and Savolainen, 2001 ) and for DNA hybridization distances for passerine birds ( Barraclough et al. , 1998 ), neither the generality of the relationship nor its underlying causes are known, so the relevance of this association to the Cambrian explosion is currently unknown.
The ideal way to test the effect of explosive radiations on the rate of molecular evolution would be to compare clades with a demonstrably rapid rate of diversification to similar lineages that have not undergone high rates of evolution. Island endemic radiations, characterized by high rates of speciation, adaptation into new ecological niches, and rapid morphological change, provide one way of making this comparison. These radiations are associated with many evolutionary processes that might effect the rate of molecular evolution: genetic bottlenecks as populations are initiated from a small number of colonists, rapid rate of phenotypic evolution as species are released from the constraints of the mainland ecosystems (such as predators), and novel adaptation as they evolve into a range of new niches. A study of thirteen island datasets, where the rate of molecular evolution for the island clades was compared to mainland relatives that showed no evidence of rapid evolution, revealed no consistent effect of rapid adaptive radiation on molecular evolution (unpublished data).
So there is currently no empirical evidence to suggest that rates of molecular evolution would have been faster in the early evolution of the metazoan kingdom. However, it is possible that some unknown effect sped the molecular clock during the Cambrian explosion—could such unknown rate variation be allowed for in molecular clock analyses? One approach is to develop molecular phylogenetic methods that explicitly model variation in substitution rate across the phylogeny as part of the phylogenetic estimation process. There are a number of new methods that incorporate variable rates, either as a random walk of rates through time ( Sanderson, 1997 ), or using a Bayesian framework ( Kishino et al. , 2001 ).
For example, Aris-Brosou and Yang (2002) used a Bayesian approach ( Thorne et al. , 1998 ; Kishino et al. , 2001 ) to allow rates of substitution of the 18S rRNA gene to vary across the phylogeny of 39 species of metazoan. Under a clock model (constant rates across the phylogeny), they estimated divergence dates for major divisions of the metazoan tree deep in the Precambrian (1,062–1,567 million years ago, Mya). But under the rate-variable model, the date estimates were dramatically younger (516–619 Mya), coinciding neatly with the appearance of the earliest multicellular animal fossils in the Vendian. It should be noted that the estimated date of the plant-animal kingdom split was also surprisingly young at around 650 Mya, in contrast to the more common age estimates of the kingdom split of at least 1,400 Mya (however the kingdom split was not a focus of the study and only one plant species was included). The authors concluded that the 18S gene underwent an acceleration in evolutionary rate in all of the sampled metazoan lineages in the early Cambrian (550–500 Mya), after which substitution rates declined across all lineages (excepting a later burst in the chordate lineage). No mechanism for this co-ordinated acceleration and deceleration of substitution rates across many independent lineages was offered ( Aris-Brosou and Yang, 2002 ).
It is important to note that, as with other phylogenetic methods, the rate-variable methods rely on a number of assumptions, some of which may be inappropriate for the data considered. For example, the Bayesian method employed by Aris-Brosou and Yang (2002) assumed a constant birth-death process over the phylogeny (lineages created by speciation and removed by extinction at an even rate throughout the history of the taxa under consideration). This presents a dilemma—although divergence date estimates around 550 Mya were interpreted to support the reality of a Cambrian explosion, the results were built on the assumption that no such explosion had occurred, because speciation rates were assumed to be constant throughout time ( Aris-Brosou and Yang, 2002 ). Species sampling strategy must also be considered: rather than being a random sample, the data used by Aris-Brosou and Yang (2002) contained representatives chosen from major metazoan lineages for which reliable fossil dates were available (see Bromham et al. , 1998 ). It is not entirely clear how to sample metazoan sequences randomly in order to satisfy the assumptions of the underlying birth-death process: if each metazoan species should have an equal chance of being included, then the dataset would probably be characterized by a preponderance of beetles. More broadly, it is currently unclear whether stochastic methods, which allow random variation in rate, would be suited to systems where rate varied systematically, for example with body size or speciation rate. Rate-variable methods, such as the Bayesian approach employed by Aris-Brosou and Yang (2002) , are a promising new approach to dating the metazoan radiation that may challenge the conclusions of many of the earlier molecular clock studies, but much work needs to be done to test their reliability.
A less sophisticated approach is to reflect the uncertainty in the molecular clock due to the possibility of temporal rate variation. For example, the observed range in substitution rates over the metazoan phylogeny can be used to put bounds on the possible divergence dates between animal phyla. For 18s rRNA and mitochondrial protein coding genes, allowing all interphylum lineages the maximum observed rate of the metazoan tree could bring the molecular dates for the deep divisions of the metazoan phylogeny closer to the Cambrian, but the date estimates were still long before the appearance of the first undisputed bilaterian fossils ( Bromham and Hendy, 2000 ). However, without a mechanism for universally faster early rates, these confidence limits are only useful to rule out some dates as being beyond reasonable inference given current understanding of rates of molecular evolution.
The discrepancy between molecular and palaeontological dates for the origin of animal phyla remains unresolved. The accuracy and precision of molecular dates should be subject to scrutiny, as there are many aspects of molecular studies that could lead to error in molecular date estimates. The wide variation between estimates shows that the precision of molecular dates leaves much to be desired. More specifically, it has been suggested that processes associated with explosive radiations—accelerated morphological change, relaxed adaptive constraint or increased speciation rate—could speed the rate of molecular evolution, making molecular date estimates for the origin of metazoan phyla too old. There is a lack of empirical evidence for this hypothesis, partly because determinants of rates of molecular evolution have not been explored for invertebrates, but also because comparative tests have thus far revealed no association between morphological evolution or adaptive radiation and rates of molecular evolution. Furthermore, rates of molecular evolution at the base of the metazoan radiation would have to have been many times higher than throughout the remainder of the Phanerozoic in order to reconcile molecular data to a Cambrian origin of the major divisions of the metazoan kingdom. It may be that future work reveals an as yet unknown mechanism for such a dramatic deceleration in molecular rates that could account for the apparently deep molecular divergence between phyla. But until such a mechanism is uncovered, it will be difficult to reconcile the molecular data to a Cambrian explosion of all animal phyla.
The bottom line is that the molecular data need to be explained, not dismissed—if the deep molecular divergences were not produced by a long Precambrian history of metazoan lineages, then they must have been produced by some other process. The imperfections of the fossil record—such as temporal gaps in preservation, taxon bias and patchy geographical representation—do not make the fossil record worthless, but they do complicate the interpretation of palaeontological data. Similarly, variation in rates of molecular evolution, imprecision of date estimates, and variation between different genes and taxa do not make the molecular data useless, but they do illustrate the danger of a simplistic interpretation of molecular distances. Molecular data must be telling us something about metazoan evolution, whether or not current molecular date estimates are correct. Much work remains to be done to arrive at an adequate explanation of the deep molecular divergence between animal phyla.
Fig. 1. Hemichordates were previously allied to chordates on the basis of the shared body plan characters of a pharynx and dorsal nerve cord. Molecular phylogenies have suggested that hemichordates are more closely related to echinoderms, a grouping that implies the deuterostome ancestor had the chordate body plan features of a pharynx and dorsal nerve cord, which were subsequently lost in the echinoderm lineage
Fig. 2. The use of nested calibration dates can lead to the use of non-independent datapoints when using regression to estimate rates of molecular evolution. In this example, each of the datapoints contains the branch a, which connects node e and species A. The inclusion of a in each datapoint results in a decrease in the degrees of freedom of the regression, and may lead to an inflation of the apparent association between time and molecular divergence
From the Symposium The Cambrian Explosion: Putting the Pieces Together presented at the Annual Meeting of the Society for Integrative and Comparative Biology, 2–6 January 2002, at Anaheim, California.
E-mail: [email protected]
I thank Andrew Rambaut for helpful discussions about rate-variable methods, and Graham Budd and Kevin Peterson for inviting me to the Cambrian Explosion symposium.
Abouheif , E. , R. Zardoya, and A. Meyer. 1998 . Limitations of metazoan 18S rRNA sequence data: Implications for reconstructing a phylogeny of the animal kingdom and inferring the reality of the Cambrian explosion. J. Mol. Evol , 47 394 -405.
Archibald , J. D. 1999 . Pruning and grafting on the mammalian phylogenetic tree. Acta Palaeontol. Pol , 44 220 -222.
Aris-Brosou , S. , and Z. Yang. 2002 . Effects of models of rate evolution on estimation of divergence dates with special reference to the metazoan 18S ribosomal RNA phylogeny. Syst. Biol , 51 703 -714.
Ayala , F. J. , A. Rzhetsky, and F. J. Ayala. 1998 . Origin of the metazoan phyla: Molecular clocks confirm palaeontological estimates. Proc. Natl. Acad. Sci. U.S.A , 95 606 -611.
Barraclough , T. G. , and V. Savolainen. 2001 . Evolutionary rates and species diversity in flowering plants. Evolution , 55 677 -683.
Barraclough , T. G. , A. P. Vogler, and P. H. Harvey. 1998 . Revealing the factors that promote speciation. Phil. Trans. R. Soc. London B , 353 241 -249.
Benton , M. J. , M. A. Willis, and R. Hitchin. 2000 . Quality of the fossil record through time. Nature , 403 534 -537.
Boore , J. L. , and J. L. Staton. 2002 . The mitochondrial genome of the sipunculid Phascolopsis gouldii supports its association with Annelida rather than Mollusca. Evolution , 19 127 -137.
Bromham , L. 2002 . Molecular clocks in reptiles: Life history influences rate of molecular evolution. Mol. Biol. Evol , 19 302 -309.
Bromham , L. 2003 . Molecular clocks and explosive radiations. J. Mol. Evol. (In press).
Bromham , L. , A. Rambaut, R. Fortey, A. Cooper, and D. Penny. 1998 . Testing the Cambrian explosion hypothesis by using a molecular dating technique. Proc. Natl. Acad. Sci. U.S.A , 95 12386 -9.
Bromham , L. , A. Rambaut, and P. H. Harvey. 1996 . Determinants of rate variation in mammalian DNA sequence evolution. J. Mol. Evol , 43 610 -21.
Bromham , L. , M. R. Q. Woolfit, M. S. Y. Lee, and A. Rambaut. 2002 . Testing the relationship between morphological and molecular rates of change along phylogenies. Evolution , 56 1921 -1930.
Bromham , L. D. , and B. M. Degnan. 1999 . Hemichordates and deuterostome evolution: Robust molecular phylogenetic support for a hemichordate+echinoderm clade. Evol. Dev , 1 166 -171.
Bromham , L. D. , and M. D. Hendy. 2000 . Can fast early rates reconcile molecular dates to the Cambrian explosion? Proc. R. Soc. London B , 267 1041 -1047.
Bromham , L. D. , A. Rambaut, M. D. Hendy, and D. Penny. 2000 . The power of relative rates tests depends on the data. J. Mol. Evol , 50 296 -301.
Budd , G. E. , and S. Jensen. 2000 . A critical reappraisal of the fossil record of the bilaterian phyla. Biol. Rev , 75 253 -295.
Carroll , S. B. 1995 . Homeotic genes and the evolution of arthropods and chordates. Nature , 376 479 -485.
Castresana , J. , G. Feldmaier-Fuchs, S. Yokobori, N. Satoh, and S. Paabo. 1998 . The mitochondrial genome of the hemichordate Balanoglossus carnosus and the evolution of deuterostome mitochondria. Genetics , 150 1115 -1123.
Conway Morris , S. 1998 . Early metazoan evolution: Reconciling paleontology and molecular biology. Amer. Zool , 38 867 -877.
Cooper , A. , and R. Fortey. 1998 . Evolutionary explosions and the phylogenetic fuse. Trends Ecol. Evol , 13 151 -156.
Cooper , A. , and D. Penny. 1997 . Mass survival of birds across the Cretaceous-Tertiary boundary: Molecular evidence. Science , 275 1109 -1113.
Darwin , C. 1859 . The origin of species by means of natural selection . John Murray, London, p. 286–289.
Doolittle , R. F. , D. F. Feng, S. Tsang, G. Cho, and E. Little. 1996 . Determining divergence times of the major kingdoms of living organisms with a protein clock. Science , 271 470 -477.
Erwin , D. H. 1993 . The origin of metazoan development: A paleobiological perspective. Biol. J. Linn. Soc , 50 255 -274.
Feng , D. F. , G. Cho, and R. F. Doolittle. 1997 . Determining divergence times with a protein clock: Update and reevaluation. Proc. Natl. Acad. Sci. U.S.A , 94 13028 -33.
Fleischer , R. C. , C. E. McIntosh, and C. L. Tarr. 1998 . Evolution on a volcanic conveyor belt: Using phylogeographic reconstructions and K-Ar based ages of the Hawaiian islands to estimate molecular evolutionary rates. Mol. Ecol , 7 533 -545.
Gillespie , J. H. 1991 . The causes of molecular evolution . Oxford University Press, Oxford.
Gu , X. 1998 . Early metazoan divergence was about 830 million years ago. J. Mol. Evol , 47 369 -371.
Haeckel , E. 1879 . The evolution of man: A popular exposition of the principal points of human ontogeny and phylogeny , London. .
Hinman , V. , and B. D. Degnan. 2000 . Retonoic acid perturbs Otx gene expression in the ascidian pharynx. Dev. Genes Evol , 210 129 -139.
Holland , P. W. H. , A. M. Hacker, and N. A. Williams. 1991 . A molecular analysis of the phylogenetic affinities of Saccoglossus cambrensis Brambell and Cole (H). Phil. Trans. R. Soc. London B , 332 185 -189.
Jenner , R. A. 2003 . Unleasing the force of cladistics? Metazoan phylgenetics and hypothesis testing. Integrative and Comparative Biology , 43 -000.
Johnson , K. P. , and J. Seger. 2001 . Elevated rates of nonsynonymous substitution in island birds. Mol. Biol. Evol , 18 874 -881.
Kimura , M. 1983 . The neutral theory of molecular evolution . Cambridge University Press, Cambridge.
Kishino , H. , J. L. Thorne, and W. J. Bruno. 2001 . Performance of a divergence time estimation method under a probabilistic model of rate evolution. Mol. Biol. Evol , 18 352 -361.
Kumar , S. , and S. B. Hedges. 1998 . A molecular timescale for vertebrate evolution. Nature , 392 917 -920.
Lee , M. S. Y. 1998 . Molecular clock calibrations and metazoan divergence dates. J. Mol. Evol , 49 385 -391.
Lee , M. S. Y. 1999 . Shortening the phylogenetic fuse. Trends Ecol. Evol , 13 323 -323.
Li , W.-H. , D. L. Ellesworth, J. Krushkal, B. H.-J. Chang, and D. Hewett-Emmett. 1996 . Rates of nucleotide substitution in primates and rodents and the generation-time effect hypothesis. Mol. Phylog. Evol , 5 182 -187.
Lyell , C. 1830 . Principles of geology . London.
Lynch , M. 1999 . The age and relationships of the major animal phyla. Evolution , 53 319 -325.
Madsen , O. , M. Scally, C. J. Douady, D. J. Kao, R. W. DeBry, R. Adkins, H. M. Amrine, M. J. Stanhope, W. W. de Jong, and M. S. Springer. 2001 . Parallel adaptive radiations in two major clades of placental mammals. Nature , 409 610 -614.
Martin , A. P. , and S. R. Palumbi. 1993 . Body size, metabolic rate, generation time and the molecular clock. Proc. Natl. Acad. Sci. U.S.A , 90 4087 -4091.
Mooers , A. Ø. , and P. H. Harvey. 1994 . Metabolic rate, generation time and the rate of molecular evolution in birds. Mol. Phylog. Evol , 3 344 -350.
Nei , M. , P. Xu, and G. Glazko. 2001 . Estimation of divergence times from multiprotein sequences for a few mammalian species and several distantly related organisms. Proc. Natl. Acad. Sci. U.S.A , 98 2497 -2502.
Ohta , T. 1993 . An examination of the generation time effect on molecular evolution. Proc. Natl. Acad. Sci. U.S.A , 90 10676 -10680.
Omland , K. E. 1997 . Correlated rates of molecular and morphological evolution. Evolution , 51 1381 -1393.
Papadopoulos , D. , D. Schneider, J. Meier-Eiss, W. Arber, R. E. Lenski, and M. Blot. 1999 . Genomic evolution during a 10,000-generation experiment with bacteria. Proc. Natl. Acad. Sci. U.S.A , 96 3807 -3812.
Penny , D. , M. Hasegawa, P. J. Waddell, and M. D. Hendy. 1999 . Mammalian evolution: Timing and implications from using the LogDeterminant transform for proteins of differing amino acid composition. Syst. Biol , 48 76 -93.
Peterson , K. J. , and D. J. Eernisse. 2001 . Animal phylogeny and the ancestry of bilaterians: Inferences from moprhology and 18s rDNA gene sequences. Evol. Dev , 3 170-2 -5.
Rambaut , A. , and L. Bromham. 1998 . Estimating divergence dates from molecular sequences. Mol. Biol. Evol , 15 442 -8.
Robinson , M. , M. Gouy, C. Gautier, and D. Mouchirod. 1998 . Sensitivity of relative rates tests to taxonomic sampling. Mol. Biol. Evol , 15 1091 -1098.
Runnegar , B. 1982 . A molecular-clock date for the origin of the animal phyla. Lethaia , 15 199 -205.
Sanderson , M. J. 1997 . A nonparametric approach to estimating divergence times in the absence of rate constancy. J. Mol. Evol , 14 1218 -1231.
Scherer , S. 1989 . The relative-rate test of the molecular clock hypothesis: A note of caution. Mol. Biol. Evol , 6 436 -441.
Simpson , G. G. 1944 . Tempo and mode in evolution . Columbia University Press, New York.
Tajima , F. 1993 . Simple methods for testing the molecular evolutionary clock hypothesis. Genetics , 135 599 -607.
Thorne , J. L. , H. Kishino, and I. S. Painter. 1998 . Estimating the rate of the rate of molecular evolution. Mol. Biol. Evol , 15 1647 -1657.
Valentine , J. W. , D. Jablonski, and D. H. Erwin. 1999 . Fossils, molecules and embryos: New perspectives on the Cambrian explosion. Development , 126 851 -859.
Wang , D. Y.-C. , S. Kumar, and S. B. Hedges. 1999 . Divergence time estimates for the early history of animal phyla and the origin of plants, animals and fungi. Proc. R. Soc. London B , 266 163 -171.
Williams , G. C. 1992 . Natural selection: Domains, levels and challenges . Oxford University Press, Oxford.
Wray , G. A. , J. S. Levington, and L. H. Shapiro. 1996 . Molecular evidence for deep Precambrian divergences among metazoan phyla. Science , 274 568 -573.
Wu , C.-I. , and W.-H. Li. 1985 . Evidence for higher rates of nucleotide substitutions in rodents than in man. Proc. Natl. Acad. Sci. U.S.A , 82 1741 -1745.
Yang , Z. H. , and R. Nielsen. 1998 . Synonymous and nonsynonymous rate variation in nuclear genes of mammals. J. Mol. Evol , 46 409 -418.
Zuckerkandl , E. , and L. Pauling. 1965 . Evolutionary divergence and convergence in proteins. In V. Bryson and H. J. Vogel (eds.), Evolving genes and proteins. Academic Press, New York.
| Month: | Total Views: |
|---|---|
| January 2017 | 5 |
| February 2017 | 25 |
| March 2017 | 13 |
| April 2017 | 10 |
| May 2017 | 4 |
| June 2017 | 2 |
| July 2017 | 6 |
| August 2017 | 7 |
| September 2017 | 9 |
| October 2017 | 14 |
| November 2017 | 24 |
| December 2017 | 21 |
| January 2018 | 26 |
| February 2018 | 35 |
| March 2018 | 34 |
| April 2018 | 52 |
| May 2018 | 22 |
| June 2018 | 21 |
| July 2018 | 15 |
| August 2018 | 20 |
| September 2018 | 24 |
| October 2018 | 31 |
| November 2018 | 42 |
| December 2018 | 24 |
| January 2019 | 18 |
| February 2019 | 31 |
| March 2019 | 72 |
| April 2019 | 33 |
| May 2019 | 32 |
| June 2019 | 22 |
| July 2019 | 20 |
| August 2019 | 48 |
| September 2019 | 48 |
| October 2019 | 63 |
| November 2019 | 62 |
| December 2019 | 39 |
| January 2020 | 23 |
| February 2020 | 38 |
| March 2020 | 38 |
| April 2020 | 59 |
| May 2020 | 27 |
| June 2020 | 29 |
| July 2020 | 29 |
| August 2020 | 36 |
| September 2020 | 45 |
| October 2020 | 34 |
| November 2020 | 42 |
| December 2020 | 81 |
| January 2021 | 15 |
| February 2021 | 69 |
| March 2021 | 104 |
| April 2021 | 61 |
| May 2021 | 61 |
| June 2021 | 29 |
| July 2021 | 11 |
| August 2021 | 18 |
| September 2021 | 32 |
| October 2021 | 29 |
| November 2021 | 64 |
| December 2021 | 29 |
| January 2022 | 45 |
| February 2022 | 58 |
| March 2022 | 49 |
| April 2022 | 46 |
| May 2022 | 35 |
| June 2022 | 28 |
| July 2022 | 16 |
| August 2022 | 21 |
| September 2022 | 36 |
| October 2022 | 31 |
| November 2022 | 26 |
| December 2022 | 40 |
| January 2023 | 28 |
| February 2023 | 36 |
| March 2023 | 48 |
| April 2023 | 31 |
| May 2023 | 9 |
| June 2023 | 20 |
| July 2023 | 9 |
| August 2023 | 12 |
| September 2023 | 12 |
| October 2023 | 46 |
| November 2023 | 40 |
| December 2023 | 24 |
| January 2024 | 17 |
| February 2024 | 31 |
| March 2024 | 24 |
| April 2024 | 38 |
| May 2024 | 23 |
| June 2024 | 34 |
| July 2024 | 28 |
| August 2024 | 26 |
| September 2024 | 16 |
Email alerts
Citing articles via.
- Recommend to your Library
Affiliations
- Online ISSN 1557-7023
- Print ISSN 1540-7063
- Copyright © 2024 The Society for Integrative and Comparative Biology
- About Oxford Academic
- Publish journals with us
- University press partners
- What we publish
- New features
- Open access
- Institutional account management
- Rights and permissions
- Get help with access
- Accessibility
- Advertising
- Media enquiries
- Oxford University Press
- Oxford Languages
- University of Oxford
Oxford University Press is a department of the University of Oxford. It furthers the University's objective of excellence in research, scholarship, and education by publishing worldwide
- Copyright © 2024 Oxford University Press
- Cookie settings
- Cookie policy
- Privacy policy
- Legal notice
This Feature Is Available To Subscribers Only
Sign In or Create an Account
This PDF is available to Subscribers Only
For full access to this pdf, sign in to an existing account, or purchase an annual subscription.

An official website of the United States government
The .gov means it’s official. Federal government websites often end in .gov or .mil. Before sharing sensitive information, make sure you’re on a federal government site.
The site is secure. The https:// ensures that you are connecting to the official website and that any information you provide is encrypted and transmitted securely.
- Publications
- Account settings
The PMC website is updating on October 15, 2024. Learn More or Try it out now .
- Advanced Search
- Journal List
- Scientific Reports
The two phases of the Cambrian Explosion
Andrey yu. zhuravlev.
1 Department of Biological Evolution, Faculty of Biology, Moscow State University named after M.V. Lomonosov, Moscow GSP-1, 119991 Russia
Rachel A. Wood
2 School of GeoSciences, University of Edinburgh, King’s Buildings, James Hutton Road, Edinburgh, EH9 3FE UK
Associated Data
The authors declare that the data supporting the findings of this study are available within the paper and its supplementary information files.
The dynamics of how metazoan phyla appeared and evolved – known as the Cambrian Explosion – remains elusive. We present a quantitative analysis of the temporal distribution (based on occurrence data of fossil species sampled in each time interval) of lophotrochozoan skeletal species (n = 430) from the terminal Ediacaran to Cambrian Stage 5 (~545 – ~505 Million years ago (Ma)) of the Siberian Platform, Russia. We use morphological traits to distinguish between stem and crown groups. Possible skeletal stem group lophophorates, brachiopods, and molluscs (n = 354) appear in the terminal Ediacaran (~542 Ma) and diversify during the early Cambrian Terreneuvian and again in Stage 2, but were devastated during the early Cambrian Stage 4 Sinsk extinction event (~513 Ma) never to recover previous diversity. Inferred crown group brachiopod and mollusc species (n = 76) do not appear until the Fortunian, ~537 Ma, radiate in the early Cambrian Stage 3 (~522 Ma), and with minimal loss of diversity at the Sinsk Event, continued to diversify into the Ordovician. The Sinsk Event also removed other probable stem groups, such as archaeocyath sponges. Notably, this diversification starts before, and extends across the Ediacaran/Cambrian boundary and the Basal Cambrian Carbon Isotope Excursion (BACE) interval (~541 to ~540 Ma), ascribed to a possible global perturbation of the carbon cycle. We therefore propose two phases of the Cambrian Explosion separated by the Sinsk extinction event, the first dominated by stem groups of phyla from the late Ediacaran, ~542 Ma, to early Cambrian stage 4, ~513 Ma, and the second marked by radiating bilaterian crown group species of phyla from ~513 Ma and extending to the Ordovician Radiation.
Introduction
The Cambrian Explosion is a phenomenon that encompasses the dramatic appearance of diverse metazoans with biomineralized skeletons, an increase in metazoan complexity and behaviour, a substrate revolution that re-organised the sedimentary record, and the development of biodiverse marine ecosystems with complex food webs 1 – 5 . The relative importance of external drivers, such as rise of oxygen or seawater chemistry changes 6 – 9 , biological drivers, such as the influence of metazoan irrigation 10 , and feedbacks between the two 11 , remains unclear. Likewise, the relationship between Ediacaran and Cambrian biotas remains unresolved, with some arguing that the Cambrian Explosion has a ‘deep root’ in the terminal Ediacaran 12 , or that the first phase of the ‘Cambrian Explosion’ was either the Nama assemblage (~550–541 Ma) 13 , or appeared even earlier at the Avalon-White Sea boundary at ~561 Ma 14 . In addition, while it has been conjectured that extinction or turnover events of metazoans occurred at ~551 Ma 13 , 15 and at the Ediacaran/Cambrian boundary at ~541 - 540 Ma (e.g. 13 , 16 ), there is no consensus as to the precise form either of these dynamics, or indeed their timing, or causes (compare 13 , 14 , 17 ).
The combined body and trace fossil record suggests the Cambrian Radiation of bilaterians may have followed a progressive two-stage diversification: the terminal Ediacaran (~560 Ma) to early Cambrian Stage 2 to 3 (mid-Tommotian to Atdabanian) interval dominated by stem groups, and after Cambrian Stage 2 to 3 when definitive crown group representatives of phyla appeared 18 . Most phylum-level body-plan evolution seems to have taken place well after the Cambrian Explosion, throughout the Cambrian and beyond; stem lineages are considered to have largely disappeared by the Ordovician 18 .
Placing extinct fossil taxa in phylogenetic order through the application of stem- and crown group concepts allows the order of character acquisition to be considered in both time and environmental context 18 . Even when highly problematic, all extinct taxa must have stem- or crown group relationships to extant taxa. A crown group is a monophyletic group consisting of the last common ancestor of all living forms and all of its descendants. A stem group is a paraphyletic group that lacks the defining morphological characters of the crown group, where all members are extinct. This therefore consists of the primitive relatives of the crown group, along the phylogenetic line up to, but not including, the last common ancestor of the crown group and their nearest living relatives 19 .
The considerable number of characters that can define crown groups were often acquired incrementally over geological time 20 . Random, background extinctions will inevitably erode the base of a clade through time, whether or not basal members are particularly prone to extinction 19 . Hence, the older a fossil, the more likely it is to fall outside the phylum-level of classification. But mass extinctions may operate quite differently, as they can remove taxa selectively based on particular ecological or other traits 21 and lead to long-lasting changes in taxonomic composition and ecosystem functioning 22 .
Here we construct a high resolution temporal distribution of skeletal species (n = 1188) from the upper Ediacaran to the basal Cambrian Series 3 of the Siberian Platform in order to understand the evolutionary dynamics of the Cambrian Explosion (see Supplementary references). The Siberian Platform formed a separate province during the Ediacaran-Cambrian 23 – 26 , where the stratigraphy and age dating is relatively well known (Fig. 1 ) and the biota diverse. New coupled high-resolution δ 13 C and biostratigraphic data as well as improved U-Pb zircon dates suggests that terminal Ediacaran – early Cambrian sections on the northern and south-eastern Siberian Platform are more complete than previously thought, and also indicate that the Cambrian Explosion as shown by the record of skeletal biota may have been a more protracted event 12 , 27 . The first diverse skeletal assemblages of Cambrian type (including various halkieriids, chancelloriids, and hyoliths in addition to anabaritids and protoconodonts), occur between levels dated from 543.9 ± 0.24 to 529.7 ± 0.3 Ma which precede the strong basal Cambrian negative carbon isotope excursion (BACE), and in some areas even the basal Terreneuvian Trichophycus pedum ichnofossil assemblage 12 , 28 – 30 . Additionally, Ediacaran shelly taxa (cloudinids) co-occur with some of the earliest Cambrian shelly taxa (anabaritids) on the south-eastern Siberian Platform, indicating a continuity of the skeletal fossil record around the Precambrian-Cambrian boundary 12 . The additional presence of late Ediacaran soft-bodied rangeomorphs, including their biomineralized holdfasts, as well as chambered palaeopascichnids and Nenoxites (= Shaanxilithes ) trace fossils found in immediately underlying strata of the same sections 12 , 27 indicates that this record is, in turn, rooted in so-called “post-Kotlinian wormworld” (e.g. 13 ).

Early Cambrian time scale for the Siberian Platform, Russia, with key radiometric dates (numbered; Siberian radiometric dates are in bold), international chronostratigraphy (ICS), and stages and zones accepted for the Siberian Platform. Radiometric dates from 1 27 , 93 ; 2 94 ; 3 95 , 4 96 ; 5 97 ; 6 98 – 100 ; 7 30 ; 8 101 . Right column shows numbered temporal units, each c. 2.5 Myr in duration. ED = Ediacaran. 3 = Cambrian Series 3, pars. Modified from 5 .
In particular we consider the distribution of stem and crown group Lophotrochozoa, which is a monophyletic clade of protostome animals within the Spiralia, consisting of Mollusca, Lophophorata, Nemertea and Annelida 31 – 33 . The Lophotrochozoa constitutes a third of all modern marine animals 34 , and was chosen as it is species-rich and represented mostly by skeletal taxa in Ediacaran-Cambrian strata. Deuterostome and cnidarian fossils are too scarce for quantitative analysis, and putative poriferans do not allow detailed character subdivision, due to either an absence of diagnostic spicules (e.g. Archaeocyatha) or the frequently disarticulated preservation of spiculate classes. More importantly, neither the temporal fossil record nor comparative characters of the Lophotrochozoa are reliant upon exceptional preservation (Lagerstätten), as has been noted in other significant groups of the radiation such as euarthropods. This taphonomic bias is exemplified by the fact that crown group euarthropods appear before (521 Ma) stem lineage euarthropods (518 Ma), due in part to differential skeletonisation 35 . Our study thus enables an understanding of how important phyla including the Mollusca, Brachiopoda and Annelida, may have been assembled, in turn informing likely selective pressures and ecological consequences.
Proposed stem-group Lophotrochozoa
The soft-bodied Ediacaran taxon Kimberella (~560 to ~550 Ma) has been proposed to represent a stem group mollusc 36 – 38 , although this placement remains problematic 17 . We exclude this from our analysis given this controversy and the lack of skeletonized hard parts.
We assign hyoliths (both hyolithimorphs and orthothecimorphs), tommotiids (including tannuolinids, Sunnaginia , and Lapworthella ), and Oymurania to stem group lophophorates, and sachitids (including halkieriids and siphogonuchitids), wiwaxiids, and, probably, maikhanellids and helcionelloids to stem group molluscs following the phylogenetic and morphological inferences detailed below.
Hyoliths (Fig. 2j ), despite their unusual, large calcareous conical shells incorporating a U-shaped intestine and an extendable tentacle-bearing lophophore, have molluscan-type microstructures, a thick compound operculum and, sometimes in hyolithimorphs, a pair of additional curved rigid lateral bar-like supports 39 , 40 .

Early and early middle Cambrian skeletal stem- ( a – f , i – j ) and crown group ( g , h ) lophotrochozoans from the Siberian Platform. ( a ) Aldanella attleborensis (Shaler & Foerste), stem mollusc, helcionelloid; shell ( 29 , Fig. 20A 1 ); ( b ) Camenella garbowskae Missarzhevsky, stem lophophorate, tommotiid; sclerite ( 102 , Fig. 37A); ( c ) Ceratoconus striatus Chen & Zhang, stem mollusc, helcionelloid; shell ( 29 , Fig. 26A 1 ); ( d ) Halkieria sp., stem lophotrochozoan; halkieriid; sclerite ( 29 , Fig. 46C 2 ); ( e ) Tannuolina pavlovi Kouchinsky et al ., stem lophophorate, tommotiid; sclerite ( 103 , Fig. 2A 2 ); ( f ) Oymurania gravestocki Ushatinskaya, stem brachiopod; valve ( 104 , Fig. 8A); ( g ) Pelmanotreta neguertchenensis (Pelman), crown brachiopod, paterinate; valve ( 105 , Fig. 2i); ( h ) Pojetaia dentifera Kouchinsky et al ., crown mollusc, bivalve; valve ( 106 , Fig. 3A); ( i ) Purella antiqua (Abaimova), stem lophotrochozoan, maikhanellid; valve ( 29 , Fig. 31B 2 ); ( j ) Khetatheca cotuiensis (Sysoev), stem lophophorate, hyolith; valve ( 29 , Fig. 50G). All photographs courtesy of Artem Kouchinsky.
While both tommotiids (Fig. 2b,e ) and halkieriids s.l . (Fig. 2d ) possess multi-element shells (scleritomes), tommotiid sclerites form a narrow conical shell and penetrated by setal canals which can preserve phosphatized setae, and exhibit dense, and fine lamination. In some cases a bivalved larval protegulum with a colleplax-plate typical of the oldest linguliformean brachiopods is present 41 – 45 . Oymurania (Fig. 2f ) has setigerous canals and two shell layers, one of which shows acrotretoid brachiopod columnar microstructure, and the other resembles the prismatic framework of paterinid brachiopods 46 . The former problematic fossil Tumulduria is now reinterpreted as a detached central portion of the ventral interarea of a paterinid brachiopod 47 .
Intact calcareous sachitid scleritomes are considered to belong to a bilateral motile organism that possessed a radula and sclerites with a branching, aesthete type of canal system found in some molluscs 48 – 50 . Complete sachitid scleritomes from the Early Ordovician are recognized as stem-group aculiferan molluscs 51 . Chancelloriid sclerites possess the same morphology and microstructure despite the presence of a markedly different scleritome of a sedentary radial-symmetrical animal 52 – 54 . Thus, a more basal position of sachitids among molluscs, or even lophotrochozoans, cannot be not excluded. Wiwaxiids, although being organic, show the same overall scleritome organization 55 .
The cup-shaped maikhanellids (Fig. 2i ) consist of merged sclerites identical to co-occurring sachitids 56 . Their cross lamellar microstructure is similar to that of some gastropods and the cap-shaped protoconch is typical of monoplacophorans 57 . Bivalved calcareous stenothecoids with their paired, serially arranged muscle scars on the inner surfaces of both valves represent a further group of mollusc-like fossils but with a set of features uncommon in crown group molluscs 54 .
The majority of Cambrian mollusc-like shells with essentially molluscan microstructures, protoconchs, and some features of torsia, are assigned to either the class Helcionelloida 58 , or subclass Archaeobranchia 59 . These mainly cup-shaped and low spiral, endogastrically coiled fossils are considered to be extinct lineages of the phylum Mollusca 58 , 59 . However, a helcionelloid affinity suggests that their untorted anatomy is due to aperture posterior emarginations, and the presence of a snorkel in some forms also suggests that helcionelloids (Fig. 2a,c ) occupy a basal position within the phylum. The archaeobranchian hypothesis also emphasises a torted basic plan and ancestral gastropod affinities 59 . A stem group rather than crown group position for helcionelloids is further supported by the presence of paired bristle-like clusters extending from the aperture of the Pelagiella shell which have a striking resemblance to the parapodial chaetae of some polychaetes 60 . Additionally, helcionelloids are characterized by a different muscle system, densely porous shells that are more common in brachiopods than molluscs, and calcitic semi-nacre microstructures which are more typical of lophophorates 61 – 63 . These observations suggest that the Helcionelloida were stem-group molluscs that retained a number of shared basal features with lophotrochozoan ancestors. Pelagiella represents the most advanced branch of helcionelloids possessing a spirally coiled shell and asymmetric muscle scars suggesting at least partial torsion 59 .
Proposed crown-group Lophotrochozoa
Early Cambrian crown-group molluscs (Fig. 2h ), are recognized among bivalves as well as gastropods of the Khairkhaniidae and Onychochilidae families, belonging to the Divasibranchia and Dextrobranchia orders, respectively 59 , 64 . Crown group Lophophorata are represented in the Cambrian by 13 orders of brachiopods (Fig. 2g ), only one of which (Lingulida) survived beyond the Palaeozoic 45 , 65 , 66 .
Quantitative temporal distribution
Total skeletal species diversity on the Siberian Platform increases from the terminal Ediacaran to the middle of Cambrian Stage 2, then declines and rises again to reach a second peak at the beginning of Stage 4, followed by an abrupt and rapid decline at the end of Stage 4, followed by recovery around the Series 2/3 boundary (Fig. 3A ). The Trilobita appears in Stage 3 and, as the most speciose group, mirrors this general trend. This is in contrast to the second most speciose group, the Archaeocyatha, which first appears in Stage 2 after which there is increase in diversity until the base of Stage 4 but then the group goes abruptly extinct shortly thereafter (Fig. 3A ).

Diversity of skeletal species through the Ediacaran – early Cambrian of the Siberian Platform. ( A ) Total diversity of all skeletal species, Trilobita, and Archaeocyatha. ( B ) Total diversity of skeletal lophotrochozoan species, and stem group and crown group representatives. Ediacaran and Cambrian chronostratigraphic subdivisions are scaled according to Fig. 1 .
Total skeletal lophotrochozoan species diversity likewise increases from the terminal Ediacaran to the middle of Cambrian Stage 2, but then declines until the middle of Stage 3, rises again to reach a second peak at beginning of Stage 4, followed by an abrupt and rapid decline until the middle of Stage 4, then followed by a further rise (Fig. 3B ).
Of these, stem group lophophorates, brachiopods, and molluscs comprise a total of 354 species, and crown-groups a total of 76 species through the Ediacaran to Cambrian Stage 5 interval. Stem lophoporates, brachiopods and molluscs (halkieriids, chancelloriids and orthothecimorph hyoliths) appeared in the terminal Ediacaran (~542.5 Ma) and show two phases of diversification: the first through the Terreneuvian, and the second during the end of Stage 3 to beginning of Stage 4 (Fig. 3B ). The first crown species are known from the late Fortunian (~537 Ma – gastropods; ~535 Ma – brachiopods and bivalves), and started to radiate later during the early Cambrian Epoch 2 (~522.5 Ma). Stem group species were devastated during the early Cambrian Stage 4 at ~513 Ma but crown group mollusc and brachiopod species, despite some changes in species composition, show no marked loss of diversity, and continued to diversify at a similar apparent rate (Fig. 3B ).
Possible taphonomic and sampling biases
Taphonomic studies have shown that the fossil record can test the proposition that marine community structure has changed over time 67 , 68 . Ediacaran to Cambrian skeletal lophotrochozoans are represented by taxa of comparable millimetric sizes, forming part of the small shelly fauna as shells and disarticulated sclerites. These fossils are generally either replaced by phosphate or present in the form of inner and outer moulds. Only lingulate brachiopods and tommotiids are preserved as original shells, and only rhynchonelliform brachiopods retain their original low-Mg calcite mineralogy. In the lower Cambrian of the Siberian Platform, such fossils are restricted to argillaceous limestones (mostly wackestones and packstones), and some grainstones, all of which accumulated onshore above either normal wave or storm wave base 69 . All fossils are extracted by the same method of dissolution in buffered acetic acid to isolate phosphatic and phosphatized shells, or moulds and steinkerns (e.g. 29 ). Worker bias is unlikely given that the assemblages reflect multiple different studies and no single worker or study dominates. We infer that taphonomic biases are minimized, and sampling biases present are shared by all small skeletal fossils.
Trends through time
Total lophotrochozoan biodiversity increases until the middle of Stage 2, but then there is a notable decline that extends to approximately the middle of Stage 3 (Fig. 3B ). This interval coincides in part with an expansion of anoxic sea floor around ~525 Ma inferred from U isotopes 70 . Stem- and crown group lophotrochozoan species show distinctly different temporal distributions, with stem group lophophorate, brachiopod and mollusc taxa originating and radiating first. The preferential extinction of stem group species in early Cambrian Stage 4, at ~513 Ma coincides with the well-known Sinsk Event, an episode of widespread shallow marine anoxia on the Siberian Platform and other locations globally, which also coincides with the major extinction of the Archaeocyatha 71 . It is probable that Archaeocyatha represent a poriferan stem group, and indeed a similar temporal separation of stem and crown group diversification is observed among other metazoans at phyla level, including the Porifera (where crown group demosponges are known by Cambrian Stage 3), Cnidaria and Echinodermata 54 , 72 – 74 .
The first probable metazoan body fossils (rangeomorphs) appeared at ~570 Ma 75 . Rangeomorphs are complex, macroscopic eukaryotes, probably stem group metazoan taxa, although an affinity higher than Porifera has been proposed 76 . Rangeomorph-dominated assemblages were devastated by the Kotlin Crisis, which marks a turnover event 15 . After this we propose two phases of the Cambrian Explosion separated by the Sinsk Event extinction. The first was dominated by non-bilaterians (Porifera, Cnidaria and Ctenophora) joined by indeterminate bilaterian stem groups at ~ 560 Ma 18 and lasted until ~513 Ma. The general increase in diversity may have been interrupted by the global expansion of anoxic sea floor around ~525 Ma. Notably, this diversification started before, and continues across, the Ediacaran/Cambrian boundary and the Basal Cambrian Carbon Isotope Excursion (BACE) interval (~541 to ~540 Ma). The BACE has been ascribed to a possible global perturbation of the carbon cycle 12 .
The second phase was marked by radiating non-bilaterian and bilaterian (here determined as brachiopod and mollusc) crown group species, and started from ~513 Ma. This second radiation phase may have been interrupted or even terminated by the late Cambrian SPICE event, which marked a further minor extinction (Fig. 4 ). Crown groups brachiopod species continued to diversify during the remainder of the Cambrian and into the Ordovician. Bivalves and gastropods also formed a significant part of total global lophotrochozoan diversity and were joined by the appearance of bryozoans and cephalopods around the Cambrian/Ordovician boundary 77 , 78 . From that time onwards their diversity remained higher than stem group lophotrochozoans, which continued to decline dramatically during the Cambrian 51 , 59 , 71 , 79 , 80 . The last stem group taxa (a few hyolith genera) went extinct in the Permian 81 .

Schematic of hypothesised non-Bilaterian (total group Porifera, Cnidaria and Ctenophora) and Bilaterian diversification during the Ediacaran-Cambrian metazoan radaition, showing the fossil record of probable earliest metazoans (shown by a rangeomorph reconstruction), the Kotlin crisis, followed by two phases of Cambrian Explosion, separated by the Sinsk Event extinction (with a possible expanded interval of anoxia during Phase 1) and extending to the Ordovician Radiation through the SPICE extinction. Non-bilaterian stem group example is a stem group archaeocyath sponge; crown group is a crown group demosponge. Bilaterian stem group is shown by a tommotiid; crown group by a trilobite.
The Sinsk Event might therefore be considered a mass extinction, which appears to have preferentially removed skeletal stem group lophotrochozoans at a point when diversity was high. This rapid removal is in contrast to background extinctions that are expected to erode the base of a clade gradually through time. We note that crown group brachiopod and mollusc do not show a marked increase in diversity after the removal of stem group lophophorate, brachiopod and mollusc taxa, but continue their former diversity trajectory. This suggests that their radiation was not dependent upon the removal of incumbent stem group taxa, but rather that crown group taxa were in some way more resilient to shallow marine anoxia or other coeval environmental perturbations. Like other mass extinctions, the Sinsk Event led to significant and long-lasting changes in taxonomic composition and ecosystems 22 , 79 , 82 .
A similar sequential faunal replacement pattern of Phanerozoic metazoans has been established in the form of evolutionary marine faunas 83 , which are in part bounded by mass extinctions. During the Ediacaran to Cambrian interval, further distinguished were the Tommotian, Cambrian s.s . and Palaeozoic faunas 84 . All these faunas were discriminated by empirical and statistical analysis of family diversity patterns only without reference to phylogenetic relationships. Their existence was, however, challenged 22 because their speciation/extinction trends could merely reflect replacement between major taxonomic groups that had coupled dynamics. But our analysis shows that evolutionary faunas may in fact be a manifestation of their composition, with the ‘Tommotian’ fauna being composed of mostly stem group lophotrophorates, molluscs and brachiopods, while the Cambrian s.s . and Palaeozoic faunas are dominated by crown group representatives of molluscs, brachiopods and many other phyla.
This pattern resembles the extinction of taxa at the Permo-Triassic boundary, when groups that originated in the early Palaeozoic either went extinct (tabulate and rugose corals, trilobites, cystoporates) or significantly declined (brachiopods, trepostomates, cryptostomates, conodonts) never to recover previous levels of diversity 85 . This is in contrast to the pattern shown by groups which appeared and diversified in the late Palaeozoic, such as gymnolaemates and new bivalve, gastropod and ammonoid orders 85 – 87 .
If ecological niches are relevant, the difference in maintaining the two phases of the Cambrian Explosion might be related to differences in ecospace that was actually “empty” for skeletal animals. During the earlier phase of stem taxa radiation (~543–513 Ma), speciation was most likely promoted by the lack of competition for existing niches. This is similar to the high rates of sympatric speciation, such as noted among modern benthic caenogastropods in lakes, where high phenotypic plasticity enables evolving ecophenotypes to diversify into different substrates (e.g. 88 ). A similar pattern of early diversification as a result of adaptations to different substrates is shown by both helcionelloid mollusc and archaeocyath sponge species during the first phase of the Cambrian Explosion inferred here. The helcionelloids underwent rapid morphogenesis 89 , and archaeocyaths display extremely high inter-habitat diversity (that is, beta-diversity) in reef communities on the Siberian Platform 90 , which may also reflect high speciation rates. Niche partitioning is not inferred, as the alpha-diversity (species number per community) remains consistently low 91 . A similar effect as a result of low competition, and also correlated with a rise in beta-diversity, has been observed to be the main driver of general diversity increase in the early Cambrian 92 . This dynamic creates the unusual situation when the boundaries of even major lower Cambrian subdivisions have not been established due to an absence of cosmopolitan species. By contrast, even though stem group diversity was significantly reduced during the later crown group brachiopod and mollusc diversification (~513–508 Ma), older niches were not completely eliminated. Thus, in the aftermath of the Sinsk extinction, crown groups were able to diversify via competition for existing niches in order to incorporate into existing communities.
Conclusions
This quantitative analysis of lophotrochozoan skeletal stem- and crown group temporal distribution suggests that the Cambrian Explosion sensu lato may be redrawn as two successive phases of morphological and functional innovation that started in the terminal Ediacaran and were separated by an extinction event. This in turn allows exploration of this phenomenon as an expansion of ecological repertoires that are tractable from the fossil record.
Methods and Data
We divide the terminal Ediacaran to Cambrian Series 2 Siberian record from ~545 to ~505 Ma based on radiometric dates into 16 temporal units based on either sub-division, or combination of one to three Siberian biostratigraphic zones to create broadly equivalent units of ~2.5 Myr each. Units start at the Ediacaran Cloudina-Namacalathus-Sinotubulites assemblage zone through transitional Ediacaran-lowermost Cambrian zones (informally named in ascending order Anabarites trisulcatus, Protohertzina anabarica , and Purella antiqua zones) through Terreneuvian and Cambrian Series 2 zones up to the basal Ovatoryctocara granulata Zone of the Cambrian Stage 5 (Series 3) (Fig. 1 ). We use the timescale for this interval from available radiometric dates from fossiliferous strata of Siberia, South China, and Avalonia 93 – 101 (see Supplementary data).
We quantify the distribution of described skeletal species (n = 1188) from the upper Ediacaran to the basal Cambrian Series 3 on the Siberian Platform (see Supplementary references). This is derived from occurrence data of fossil taxa sampled in each time interval. In particular we quantify the temporal distribution of lophotrochozoan skeletal species (n = 430) (see Supplementary data). Chancelloriids, although they may belong to stem lophotrochozoans, are excluded from analysis due to their frequently disarticulated nature.
Electronic supplementary material
Acknowledgements.
Artem Kouchinsky is thanked for kindly providing all photographic images, and Jen Hoyal Cuthill, Doug Erwin, and an anonymous reviewer for thoughtful reviews.
Author Contributions
A.Yu. Z. collated the data and A.Yu. Z. and R.W. designed the research, wrote the main manuscript text, and prepared the figures.
Data Availability
Competing interests.
The authors declare no competing interests.
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information accompanies this paper at 10.1038/s41598-018-34962-y.
COMMENTS
The Cambrian explosion is a long standing macroevolutionary issue, which has been puzzling paleontologists and evolutionary biologists since 1830s as Conway Morris (2000, p. 4426) stated "William Buckland knew about it, Charles Darwin characteristically agonized over it, and still we do not fully understand it." It is generally accepted that essentially all of the readily fossilizable ...
The Cambrian explosion, as it is called, produced arthropods with legs and compound eyes, worms with feathery gills and swift predators that could crush prey in tooth-rimmed jaws. Biologists have ...
The Cambrian Explosion by nature is a three-phased explosion of animal body plans alongside episodic biomineralization, pulsed change of generic diversity, body size variation, and progressive increase of ecosystem complexity. The Cambrian was a time of crown groups nested by numbers of stem groups with a high-rank taxonomy of Linnaean system (classes and above). Some stem groups temporarily ...
The Cambrian explosion (also known as Cambrian radiation [1] or Cambrian diversification) is an interval of time approximately in the Cambrian period of the early Paleozoic when a sudden radiation of complex life occurred, and practically all major animal phyla started appearing in the fossil record. [2] [3] [4] It lasted for about 13 [5] [6] [7] to 25 [8] [9] million years and resulted in the ...
The Cambrian Explosion is a phenomenon that encompasses the dramatic appearance of diverse metazoans with biomineralized skeletons, an increase in metazoan complexity and behaviour, a substrate ...
The Cambrian-explosion hypothesis claims that this fantastic animal menagerie diverged from a common ancestor and become a recognizable set of body plans in a mere 20 million years or so. The earliest Cambrian—marked by burrows and small, strange, shelly fossils—culminates in a spectacular array of forms by about 520 Ma.
The Cambrian explosion, as it is called, produced arthropods with legs and compound eyes, worms with feathery gills and swift predators that could crush prey in tooth-rimmed jaws. Biologists have ...
Recent hypotheses for the Cambrian explosion fall into three main categories: developmental/genetic, ecologic, and abiotic/environmental, with geochemical hypotheses forming an abundant and distinctive subset of the last . Most of these hypotheses have been posited as stand-alone processes that were the main cause of the explosion, yet many of ...
The Cambrian Explosion is a polythetic event in natural history and manifested in many aspects. No simple, single cause can explain the entire phenomenon. Diagram reminiscent of eruptive evolution ...
The Cambrian 'explosion' is widely regarded as one of the fulcrum points in the history of life, yet its origins and causes remain deeply controversial. New data from the fossil record, especially of Burgess Shale-type Lagerstätten, indicate, however, that the assembly of bodyplans is not only largely a Cambrian phenomenon, but can already ...
Ediacaran Cambrian 580 Myr: Large Ediacaran animals appeared. Big animals emerged during the Ediacaran period, but these creatures were slow or immobile. A rise in oceanic oxygen concentrations at the end of the period might have helped to trigger the Cambrian evolutionary explosion. 800 600 500 400 When life sped up? 542 Myr: Extinction of
hypothesis implied that the Cambrian Explosion was accom-panied with a major extinction event, which re-established the importance of the Cambrian Explosion (Seilacher 1997). Now most Ediacaran workers tend to consider Ediacarans as a grab bag of disparate life forms (Watson 2020) rather than treat them as a homogeneous group. A couple of vendobi-
The early history of the Metazoa, whether originating as part of a Cambrian "explosion" or with an extended, Precambrian "phylogenetic fuse," remains controversial (1-3).The Cambrian explosion hypothesis—that the phyla and even classes of the animal kingdom originated in a rapid evolutionary radiation at the base of the Cambrian at 545 million yr ago (Ma) or 10-15 Ma before this ...
Ordovician radiation. Cambrian explosion, the unparalleled emergence of organisms between 541 million and approximately 530 million years ago at the beginning of the Cambrian Period. The event was characterized by the appearance of many of the major phyla (between 20 and 35) that make up modern animal life. Many other phyla also evolved during ...
Summary. The sudden appearance of fossils that marks the so-called 'Cambrian explosion' has intrigued and exercised biologists since Darwin's time. In On the Origin of Species, Darwin made it clear that he believed that ancestral forms 'lived long before' their first fossil representatives. While he considered such an invisible record ...
Our results therefore offer a new hypothesis for the timing and origin of biomineralization and the Cambrian explosion, both of which lag by tens of millions of years the initial origin of ...
Abstract. Abrupt appearance of major bilaterian clades in the fossil record during the first three stages of the Cambrian Period has puzzled the scientific world since 1830s. Many proposed causes including environmental, developmental, and ecological hypotheses, are reviewed. Nutrient availability, oxygenation, and change of seawater ...
Each hypothesis outlined here is a via-ble mechanism for increasing mean spe-cies diversity within habitat, differentiation ... diversity. However, it is unlikely that any single casual mechanism can explain the Cambrian explosion, with many of the indi-vidual hypotheses instead acting as compo-nents of interacting feedback loops between Earth ...
The Cambrian 'explosion' is widely regarded as one of the fulcrum points in the history of life, yet its origins and causes remain deeply controversial. ... This hypothesis supposes that the 'invention' of the 'set-aside cells' was subsequently utilized to allow the abrupt evolution of macroscopic adult forms which initiated not the ...
Evidence of changes in the seawater chemistry is captured in the rock record by high rates of carbonate mineral formation early in the Cambrian, as well as the occurrence of extensive beds of glauconite, a potassium-, silica-, and iron-rich mineral that is much rarer today. The influx of ions to the oceans also likely posed a challenge to the ...
The hypothesis that body plans were formed in the Cambrian Explosion then canalized predicts that there are critical characters that not only define phyla, but were instrumental in their formation (e.g., Carroll, 1995; Erwin, 1993; Valentine et al., 1999).
Introduction. The Cambrian Explosion is a phenomenon that encompasses the dramatic appearance of diverse metazoans with biomineralized skeletons, an increase in metazoan complexity and behaviour, a substrate revolution that re-organised the sedimentary record, and the development of biodiverse marine ecosystems with complex food webs 1-5.The relative importance of external drivers, such as ...
Discredited hypotheses for the Cambrian explosion. As understanding of the events of the Cambrian becomes clearer, data have accumulated to make some postulated causes for the Cambrian explosion look improbable. Some examples are the evolution of herbivory, vast changes in plate tectonic rates or orbital motion, or different evolutionary ...